|
PIT00322 Неменделевская родословная В нашей модели CellBook определение наследования как перемешивания клеточных линий можно рассмотреть на интересном примере, взятом из недавней статьи {Besseling2016} Judith Besseling, and Henrik Bringmann. Engineered non-Mendelian inheritance of entire parental genomes in C. elegans // Nature Biotechnology (2016). doi 10.1038/nbt.3643 в которой искусственно создано неменделевское наследование всего материнского или отцовского генома в Caenorhabditis elegans путем изменения структуры митотического веретена во время первого клеточного деления зиготы. Использование зародышевой специфической сверх экспрессии одного белка, сохраняющегося силовым регулятором микротрубочек GPR-1 [см ниже Redemann2011], позволило увеличивать силы, которые тянут полюсами веретена для преобразования одного биполярного митотического шпинделя до двух монопольных шпинделей. Это порождает два клеточных эмбриона, в которых одна клетка содержит только материнские хромосомы, а другая клетка содержит только отцовские хромосомы. В результате этого по мере развития эмбриона, каждая клетка животного, в том числе и зародышевые клетки, содержит генетический материал только одного из родителей в гибридном животном Такую инженерию неменделевского наследования удалось сделать благодаря результатам статьи {Redemann2011} Stefanie Redemann, Siegfried Schloissnig, Susanne Ernst, Andrey Pozniakowsky, Swathi Ayloo, Antony A. Hyman, and Henrik Bringmann. Codon adaptation-based control of protein expression in C. elegans // Nature methods 8 (3), 250-252 (2011). doi 10.1038/nmeth.1565 в которой представлен способ контролировать уровень белка нативной генетической регуляцией в Caenorhabditis elegans с помощью синтетических генов с адаптированными кодонами. Было обнаружено, что сила, действующая на шпинделе в C. elegans эмбрионов, связана с количеством регулятора G-белка GPR-1/2. Кодон-адаптированные версии любого C. elegans гена можно сконструировать с помощью веб-инструмента, или адаптера кодонов C. elegans, созданного авторами статьи [Redemann2011]. Яркое биологическое применение описано в недавней статье Основные моменты статьи Графическое резюме статьи 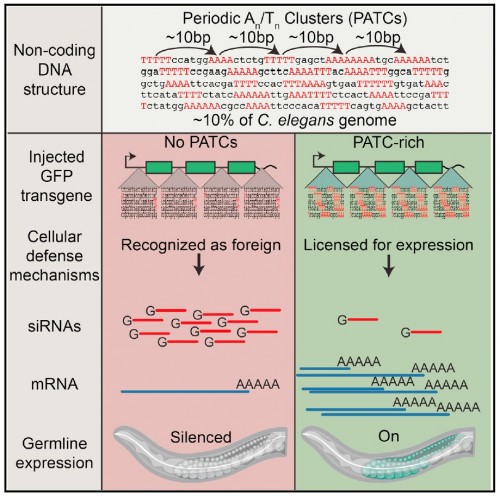 Некодирующие ДНК могут подавить стохастическое замалчивание генов в родословной (Germline) C. elegans. Клетки извлекают пользу от "глушения=замалчивания" чужеродных генетических элементов, но должны одновременно избегать инактивации собственных генов. Хотя модификации хроматина и РНК способствуют поддержанию молчащих состояний, создание молчащих областей неизбежно отражает основную последовательность и/или структуру ДНК. Показано, что распространяющаяся некодирующая особенность ДНК в Caenorhabditis elegans, характеризуется 10 парами оснований периодических Чем отличается менделевское и неменделевское наследования хорошо написал {Perkel2016} Jeffrey M. Perkel. Engineering Sexual Reproduction, Peter Belobrov 08 Sep 2016 10:38
© International Open Laboratory for Advanced Science and Technology — MOLPIT, 2009–2026
|
