|
PIT00677 Нерешаемые проблемы биофизики Решаемые н нерешаемые проблемы Книга Льва Александровича [1] классическая, где обсуждаются проблемы биологической физики, которые в традиционных учебниках обходят стороной. Каждый биофизик обязан периодически почитывать эту книгу, чтобы не впасть в распространённые заблуждения при представлении биологических переменных на языках других наук. В качестве примера одной из нерешаемых проблем, на наш взгляд, является противоречие между познанием и когнитивными науками PIT00676, созданными для обслуживания ИИ. 1. Блюменфельд Лев Александрович. Решаемые н нерешаемые проблемы биологической физики. М.: Едиториал УРСС, 2002. 160 с. Peter Belobrov 12 Jul 2026 13:14 "Although cognition is poznanie, but AI doesn't cognition anything!" «Хотя cognition — это познание, но ИИ ничего не познает!» Познающие — не когнитивные! Познание — не когниция! ИИ ничего не познаёт! Эти аксиомы давно забыты. Посему все чувства, образы и мысли давно забыты, давно забиты мусором ИИ + битовое потребление практически так оцифровано, что любовь увы забыта для когнитивных функций. Впрочем, почти 200 лет назад Фёдор Иванович Тютчев про это сказал так: полезно посмотреть PIT00608. 10 лет назад была сделана попытка введения «полной когнитивной разгрузки» [1] с экспоненциальным падением цитируемости. Снижение значения когнитивной разгрузки связано с пониманием важности истинного познания в современную эпоху, которое возникает после прочтения всего сборника [2], изданного Институтом когнитивных наук. 4 дня назад опубликован препринт [3], в котором сделана попытка представить когнитивную разгрузку в качестве универсальной для живых и искусственных систем. Когнитивная разгрузка — это когнитивная универсальность [3]. Чтобы серьёзно понимать суть противоречия между познанием и когнитивностью, полезно прочесть [4]. Эта книга написана на основе лекций на философском факультете МГУ. Ссылка взята из PIT00658. Лекции «Критика искусственного разума» Философский факультет МГУ Человечеству сегодня нужно ответить на вопрос: сможет ли оно пережить новый неолит? Пройти вторую антропологическую развилку? Если первый неолит отделил земледелие от охоты и собирательства и направил нас по пути цивилизации, то новый неолит разделяет разум на сознательный ум и искусственный. В курсе лекций разъясняется, чем искусственный интеллект отличается от органического, а органический – от человеческого. Из курса лекций студенты узнают, что все живое знает пределы своей формы; нарушение пределов формы стало причиной существования сознания; пределом формы жизни являются видимости, которые в Московской антропологической школе называют трансцендентальными галлюцинациями. На самом деле ситуация далеко не простая! По-видимому, она является неразрешимой проблемой биологической физики, чему будет посвящён следующий пост PIT00677. 1. Risko E. F., Gilbert S. J. Cognitive offloading // Trends in cognitive sciences. – 2016. – Т. 20. – №. 9. – С. 676-688. doi 10.1016/j.tics.2016.07.002. Cited 2047. Риско Э. Ф., Гилберт С. Дж. Когнитивная разгрузка. 2. Rebecca F. Schwarzlose (ed). Cognition in the Modern Era // Trends in Cognitive Sciences (TiCS), 21 (2), 57-150 (2017). Специальный выпуск «Познание в современную эпоху». 3. Fields C., Levin M. Cognitive Offloading Is a Cognitive Universal // Preprint 2026. Posted: 08 July 2026. doi 10.20944/preprints202607.0507.v1. Филдс К., Левин М. Когнитивная разгрузка — это когнитивная универсальность. Аннотация. Люди обычно переносят когнитивные задачи в окружающую среду. Здесь мы, используя только базовые принципы физики и принцип свободной энергии, показываем, что все системы обработки информации, сохраняющие свои свойства во времени, переносят задачи обработки информации в окружающую среду. Следовательно, все когнитивные системы участвуют в когнитивной перегрузке. Мы показываем, как построение экологической ниши, кинематическая репликация, биоэлектрическая сигнализация, разработка коммуникационных систем на основе общей семантики и способность LLM (Large Language Model = больших языковых моделей) демонстрировать беглое использование языка в отсутствие внеязыкового ввода — все это иллюстрирует этот процесс перегрузки. Мы заключаем, что как теоретическое понимание способностей к решению проблем, так и разработка таких способностей в артефактах будут улучшены за счет рассмотрения активных вычислений окружающей средой как повсеместного дополнения к познанию как в живых, так и в искусственных системах. Keywords: bioelectricity; complexity; efficiency; free energy principle; information flow; LLMs; morphogenesis; niche construction; shared semantics. Ключевые слова: биоэлектричество; сложность; эффективность; принцип свободной энергии; поток информации; LLMs; морфогенез; нишевое строительство; общая семантика. 4. Гиренок Ф.И. Критика искусственного разума. Монография. Изд-во «Кучково поле». 2026. 224 с. Peter Belobrov 12 Jul 2026 02:00 PIT00675 "Cochlea-on-a-Chip" «улитка-на-чипе» За 10 лет разработка «улитки-на-чипе» сильно продвинулось, правда, на русском статей нет. 1. Shen, Tian, Shanying Han, Weiwei He, Wen Yang, Xinghua Tang, Xiaolong Zhao, Xinghong Liu et al. Advances in Microfluidic Cochlea‐On‐A‐Chip // Advanced Science 12 (29), 2406077 (2025). doi 10.1002/advs.202406077. Шэнь, Тянь, Шаньин Хань, Вэйвэй Хэ, Вэнь Ян, Синхуа Тан, Сяолун Чжао, Синхун Лю и др. Достижения в области микрофлюидной системы «улитка-на-чипе». 2. Chang, Jinke, Sita Tarini Clark, Iwan Roberts, Filip Hrncirik, Zhipeng Zhang, and Manohar Bance. Artificial hearing systems based on functional cochlea models // International Journal of Extreme Manufacturing 7 (1), 012003 (2025). doi 10.1088/2631-7990/ad8aa9. Чанг, Джинке, Сита Тарини Кларк, Иван Робертс, Филип Хрнчирик, Чжипэн Чжан и Манохар Банс. Искусственные системы слуха на основе функциональных моделей улитки. Peter Belobrov 11 Jul 2026 14:30 PIT00674 Биологическая микрофлюидика улитки Почти десятилетие назад пост PIT00305 кратко описывал проблему биологической микрофлюидики ушной улитки, хотя устройство внутреннего уха так хитроумно решает проблему звукового распознавания, что пришлось вернуться и к анатомии, и к точному пониманию движения жидкостей в улитке. Без прослушивания лекции об анатомии улитки внутреннего уха приблизиться к пониманию нельзя. Так что, потратьте 20 - 30 минут, слушая Эдгара Кафарова, конечно, с карандашиком над рисунком с улиткой ниже с поста PIT00305. Кстати, после разбирательства рисунок пришлось переделывать, и уточнять подпись к рисунку. 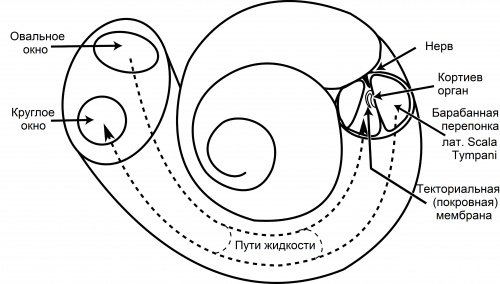 Дополнительные пояснения. Барабанная перепонка (лат. Scala Tympani), улитковый проток или барабанная лестница это, по определению, нижний костный проход улитки. Текториальная мембрана является частью кортиева органа во внутреннем ухе. Это довольно жесткая конструкция, к которой присоединяются реснички волосковых клеток. Базилярная мембрана (Basilar Membrane) - мембрана в улитке внутреннего уха, разделяющая два из трех каналов (лестниц), которые тянутся вдоль всей длины спирали улитки (улитковый проток и барабанную лестницу). Peter Belobrov 11 Jul 2026 13:26 PIT00673 Классики открытия множества полов На лекции в Свердловске в 1964 году Тимофеев-Ресовский говорил «Мы еще до конца не понимаем, в строгой и ясной форме не показали, почему на нашей планете эволюция живых форм привела именно к дву-, а не к трех- или пятиполовости. Потому что мы знаем о существовании многополовости, в частности у ряда одноклеточных водорослей. Давно и в точной форме, в основном школами Книпа и Хартмана обнаружены виды, у которых имеется более десятка полов». Эти слова мы привели 10 лет назад в посте PIT00356, но не обратили внимания на классиков открытия множества полов – Ханса Книпа – Karl Johannes Kniep и Макса Хартмана – Max Hartmann. Сейчас в открытом доступе доступно академическое издание лекций Николая Владимировича [1], которое полнее, и также как в PIT00356, его можно скачать. Карл Йоханнес Книп, известный как Ханс (3/4/1881 – 17/11/1930), был немецким ботаником и университетским профессором. Его официальное ботаническое имя — „Kniep“ = «Книп». Жизнь. Ханс Книп изучал медицину в Кильском университете, затем ботанику в Йене под руководством Эрнста Шталя, где в 1904 году получил докторскую степень. Во время учебы в Йене он также учился у Роберта Ходата в Женевском университете. С 1905 года он работал ассистентом Вильгельма Пфеффера в Лейпцигском университете, а также совершил длительную исследовательскую поездку в Берген (Норвегия), где занимался изучением биологии прорастания и физиологии водорослей (Fucus). С 1907 года он был частным лектором во Фрайбургском университете. В 1911 году Книп был назначен доцентом Страсбургского университета, а в 1914 году — профессором Вюрцбургского университета, где он занимал должность декана, а с 1923 по 1924 год — ректора. С 1924 года (после Готлиба Хаберландта) он работал профессором физиологии растений в Берлинском университете. В 1923 году он был избран членом Леопольдинского общества. Области исследований. В своих работах Книп уделял основное внимание физиологии растений, в частности хемотаксису, фотосинтезу и питанию морских растений, а также половому размножению низших растений. Его работы по цитологии и генетике грибов, особенно базидиомицетов, имеют особое значение. Он исследовал, среди прочего, образование парных ядер у гименомицетов и условия образования перемычек у базидиомицетов. С 1916 года он был редактором журнала по ботанике (der Zeitschrift für Botanik). В 1928 году издана книга [2]. Макс(имилиан) Хартманн (7/6/1876 – 11/10/1962) — зоолог и естествоиспытатель, университетский профессор и директор Института биологии им. Кайзера Вильгельма. Жизнь. Он начал обучение в Ашаффенбургской лесной академии, вскоре переключившись на зоологию и естественные науки в Мюнхенском университете, где в 1901 году получил докторскую степень под руководством Рихарда фон Хертвига. После работы ассистентом в Зоологическом институте Гисенского университета в 1902 году он получил квалификацию университетского преподавателя в 1903 году, защитив работу «Репродуктивные способы организмов». Затем он перешел в Берлинский институт инфекционных заболеваний, который позже стал Институтом Роберта Коха, и с 1909 года работал там профессором. С 1914 года он был директором Института биологии им. Кайзера Вильгельма в Берлине. Будучи председателем «Биологическо-медицинской секции» Общества им. Кайзера Вильгельма (KWG), Хартманн также был членом Сената KWG с 1929 по 1932 год. Его особые научные интересы включали физиологию оплодотворения и полового размножения, результаты опубликованы в [3, 4]. Основная литература 1. 2. Kniep, Hans. Die Sexualität der niederen Pflanzen: Differenzierung, Verteilung, Bestimmung und Vererbung des Geschlechts bei den Thallophyten (Половое поведение низших растений: дифференциация, распространение, определение и наследование пола у таллофитов). G. Fischer, 1928. 3. Hartmann, Max. Die Sexualität der Protisten und Thallophyten und ihre Bedeutung für eine allgemeine Sexualitätstheorie (Половое поведение протистов и таллофитов и его значение для общей теории полового поведения). // "Das Handbuch der Vererbungswissenschaft" («Справочник по наследственности») p.76-126 (1930). 4. Hartmann, Max. Die Sexualität: Das Wesen und die Grundsetzlichkeiten des Geschlechts und der Geschlechtsbestimmung im Tier und Pflanzenreich (Половое поведение: Природа и основы пола, и определения пола в царствах животных и растений). 1st ed. 1943. 2nd ed. 1956. Peter Belobrov 10 Jul 2026 23:18
© International Open Laboratory for Advanced Science and Technology — MOLPIT, 2009–2026
|
