|
PIT00673 Классики открытия множества полов На лекции в Свердловске в 1964 году Тимофеев-Ресовский говорил «Мы еще до конца не понимаем, в строгой и ясной форме не показали, почему на нашей планете эволюция живых форм привела именно к дву-, а не к трех- или пятиполовости. Потому что мы знаем о существовании многополовости, в частности у ряда одноклеточных водорослей. Давно и в точной форме, в основном школами Книпа и Хартмана обнаружены виды, у которых имеется более десятка полов». Эти слова мы привели 10 лет назад в посте PIT00356, но не обратили внимания на классиков открытия множества полов – Ханса Книпа – Karl Johannes Kniep и Макса Хартмана – Max Hartmann. Сейчас в открытом доступе доступно академическое издание лекций Николая Владимировича [1], которое полнее, и также как в PIT00356, его можно скачать. Карл Йоханнес Книп, известный как Ханс (3/4/1881 – 17/11/1930), был немецким ботаником и университетским профессором. Его официальное ботаническое имя — „Kniep“ = «Книп». Жизнь. Ханс Книп изучал медицину в Кильском университете, затем ботанику в Йене под руководством Эрнста Шталя, где в 1904 году получил докторскую степень. Во время учебы в Йене он также учился у Роберта Ходата в Женевском университете. С 1905 года он работал ассистентом Вильгельма Пфеффера в Лейпцигском университете, а также совершил длительную исследовательскую поездку в Берген (Норвегия), где занимался изучением биологии прорастания и физиологии водорослей (Fucus). С 1907 года он был частным лектором во Фрайбургском университете. В 1911 году Книп был назначен доцентом Страсбургского университета, а в 1914 году — профессором Вюрцбургского университета, где он занимал должность декана, а с 1923 по 1924 год — ректора. С 1924 года (после Готлиба Хаберландта) он работал профессором физиологии растений в Берлинском университете. В 1923 году он был избран членом Леопольдинского общества. Области исследований. В своих работах Книп уделял основное внимание физиологии растений, в частности хемотаксису, фотосинтезу и питанию морских растений, а также половому размножению низших растений. Его работы по цитологии и генетике грибов, особенно базидиомицетов, имеют особое значение. Он исследовал, среди прочего, образование парных ядер у гименомицетов и условия образования перемычек у базидиомицетов. С 1916 года он был редактором журнала по ботанике (der Zeitschrift für Botanik). В 1928 году издана книга [2]. Макс(имилиан) Хартманн (7/6/1876 – 11/10/1962) — зоолог и естествоиспытатель, университетский профессор и директор Института биологии им. Кайзера Вильгельма. Жизнь. Он начал обучение в Ашаффенбургской лесной академии, вскоре переключившись на зоологию и естественные науки в Мюнхенском университете, где в 1901 году получил докторскую степень под руководством Рихарда фон Хертвига. После работы ассистентом в Зоологическом институте Гисенского университета в 1902 году он получил квалификацию университетского преподавателя в 1903 году, защитив работу «Репродуктивные способы организмов». Затем он перешел в Берлинский институт инфекционных заболеваний, который позже стал Институтом Роберта Коха, и с 1909 года работал там профессором. С 1914 года он был директором Института биологии им. Кайзера Вильгельма в Берлине. Будучи председателем «Биологическо-медицинской секции» Общества им. Кайзера Вильгельма (KWG), Хартманн также был членом Сената KWG с 1929 по 1932 год. Его особые научные интересы включали физиологию оплодотворения и полового размножения, результаты опубликованы в [3, 4]. Основная литература 1. 2. Kniep, Hans. Die Sexualität der niederen Pflanzen: Differenzierung, Verteilung, Bestimmung und Vererbung des Geschlechts bei den Thallophyten (Половое поведение низших растений: дифференциация, распространение, определение и наследование пола у таллофитов). G. Fischer, 1928. 3. Hartmann, Max. Die Sexualität der Protisten und Thallophyten und ihre Bedeutung für eine allgemeine Sexualitätstheorie (Половое поведение протистов и таллофитов и его значение для общей теории полового поведения). // "Das Handbuch der Vererbungswissenschaft" («Справочник по наследственности») p.76-126 (1930). 4. Hartmann, Max. Die Sexualität: Das Wesen und die Grundsetzlichkeiten des Geschlechts und der Geschlechtsbestimmung im Tier und Pflanzenreich (Половое поведение: Природа и основы пола, и определения пола в царствах животных и растений). 1st ed. 1943. 2nd ed. 1956. Peter Belobrov 10 Jul 2026 23:18 «Extracellular Vesicle» Volume 7 (In progress). 1. «Advancements in extracellular vesicle research Достижения в исследованиях внеклеточных везикул». Bingrui Li, Valerie S. Kalluri, Alissa M. Weaver at al. Статья 100107 Обзор. 2. «Hype versus reality of artificial intelligence (AI) platforms: unmasking the limitations of large language models in the use of scientific writing and reporting Шум и реальность платформ искусственного интеллекта (ИИ): выявление ограничений больших языковых моделей в использовании в научном письме и отчетности». Bingrui Li, Ke Cheng, Raghu Kalluri. Статья 100106. doi 10.1016/j.vesic.2026.100106 3. «Injected extracellular vesicles and other nanoparticles hitchhike on erythrocytes and platelets from circulation towards organ clearance Введенные внеклеточные везикулы и другие наночастицы перемещаются из кровообращения в органы вместе с эритроцитами и тромбоцитами. Svetlana Pavlova, Doste R. Mamand, Daniel W. Hagey at al. Статья 100103. doi 10.1016/j.vesic.2025.100103 4. «Mitochondria-containing large extracellular vesicles target mouse motor neurons upon intramuscular injection Содержащие митохондрии крупные внеклеточные везикулы при внутримышечной инъекции воздействуют на двигательные нейроны мышей». Paromita Paul Pinky, Zhong-Min Wang, Purva Khare at al. Статья 100100. doi 10.1016/j.vesic.2025.100100 5. «Systematic characterization of mammalian extracellular vesicles using nano-flow cytometry Систематическая характеристика внеклеточных везикул млекопитающих с использованием нанопроточной цитометрии». Benjamin T. Vyzourek, Dirk Anderson, Luke Skrabal at al. Статья 100098. doi 10.1016/j.vesic.2025.100098 6. «Exofection by exosomes: A transient functional cargo transfer Экзофекцию с помощью экзосом: временный функциональный перенос груза». Ramkumar Menon, Madhuri Tatiparthy, Jessica Selim at al. Статья 100081. doi 10.1016/j.vesic.2025.100081 7. «Plasma extracellular vesicle cargo microRNAs are associated with heart failure and cardiovascular death following acute coronary syndrome МикроРНК, содержащиеся в внеклеточных везикулах плазмы, связаны с сердечной недостаточностью и сердечно-сосудистой смертью после острого коронарного синдрома». Worawan B. Limpitikul, Michael G. Silverman, Nedyalka Valkov at al. Статья 100070. doi 10.1016/j.vesic.2025.100070 See more at Extracellular Vesicle | Vol 7, In progress (June 2026) Peter Belobrov 19 Mar 2026 20:52 Это продолжение поста PIT00557 "Капли жизни: биологические LLPS", но здесь проявится двойственность основного закона жизни - гармония самосозидания и саморазрушения иерархии капель, клякс и клеток в коллективных совместных действиях. Peter Belobrov 18 Mar 2026 02:16 PIT00649 Одноклеточные технологии родословных 7 лет спустя при обновлении PIT00471 возникла необходимость в написании этого поста. Дело в том, что Wagner, Daniel E., and Allon M. Klein. Lineage tracing meets single-cell omics: opportunities and challenges (Отслеживание родословных встречает одноклеточные омикс-технологии: возможности и проблемы) // Nature Reviews Genetics 21 (7), 410-427 (2020). doi 10.1038/s41576-020-0223-2; Cited by 649. Baysoy, Alev, Zhiliang Bai, Rahul Satija, and Rong Fan. The technological landscape and applications of single-cell multi-omics (Технологический ландшафт и области применения мультиомиксных исследований отдельных клеток) // Nature Reviews Molecular Cell Biology 24, no. 10 (2023): 695-713. doi 10.1038/s41580-023-00615-w; Cited by 964. Yildiz, Umut, Sara Lobato-Moreno, Annique Claringbould, Hanke Gwendolyn Bauersachs, Nila H. Servaas, Evi P. Vlachou, Christian Arnold et al. "Single-cell ultra-high-throughput multiplexed chromatin accessibility and gene expression sequencing (SUM-seq) (Высокопроизводительное мультиплексное секвенирование доступности хроматина и экспрессии генов в отдельных клетках (SUM-seq)) // Nature Protocols (2026): 1-37. doi 10.1038/s41596-025-01310-0. Peter Belobrov 15 Mar 2026 14:28 Количественные и качественные категории, формальная и содержательная классификация - это разные характеристики образа осмысленных измерений живыми существами. Утром была записана идея решения метрологических проблем с содержательными образными переменными и методов действия в этих научных переменных. Требуется глубокое обдумывание и поиск формул для представления их на языке таких переменных. 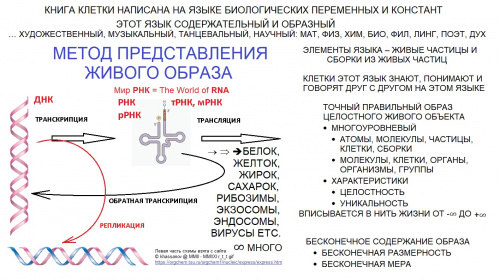 Образ представления жизни = Вселенная жизни или Жизнь Вселенной История этого поста (из Comments) #1 by PIT 18 Oct 2024 10:24 #2 by PIT 18 Oct 2024 10:54 #3 by PIT 25 Oct 2024 00:34 Peter Belobrov 17 Oct 2024 10:54 PIT00557 Капли жизни: биологические LLPS Прогрессу в понимании роли LLPS в организации капельных органелл в клетке PIT00363 до капель жизни, или просто живых капель, мы обязаны Владимиру Николаевичу Уверскому. Он один из первых увидел в денатурированных белках основную сущность внутренне неупорядоченных молекул, которые составляют большинство гибких структур в живой клетке. Поэтому новая книга про капли жизни под редакцией В.Н. Уверского получилась весьма содержательной. Droplets of Life: Membrane-Less Organelles, Biomolecular Condensates, and Biological Liquid-Liquid Phase Separation. Edited by: Vladimir N. Uversky. Academic Press, 2023. 720 p.  О книге. Капли жизни: безмембранные органеллы, биомолекулярные конденсаты и разделение биологических фаз жидкость-жидкость (LLPS) содержат фундаментальную информацию о биофизике, биогенезе, структуре, функциях и роли безмембранных органелл. Изучение фазового разделения жидкость-жидкость привлекло большое внимание таких дисциплин, как клеточная биология, биофизика, биохимия и других, пытающихся понять, как, почему и какую роль эти конденсаты играют в гомеостазе и болезненных состояниях живых организмов. Редактор этой книги нанял группу международных экспертов, чтобы предоставить актуальный и авторитетный обзор всех аспектов, связанных с этой захватывающей областью. Ключевые особенности Часть I: Введение в безмембранные органеллы и биомолекулярные конденсаты Часть II: Биология безмембранных органелл Часть III: Патологические роли LLPS Peter Belobrov 08 Jan 2023 01:24 PIT00548 Луиджи Серафини в 4K Сейчас, когда образные или картиночные меры, которыми можно проводить измерение биологических переменных, становятся уже достаточно понятными в определённых кругах исследователей, важно познакомиться с разрешением 4K - обозначение разрешающей способности в компьютерной графике, примерно соответствующее 4000 пикселей по горизонтали. Рождение 4К видеокамеры FDR-AX100 впечатляет гораздо больше, чем развитие качества видео от DVD до Blu-ray. Конечно же цифровому разрешению непостижимо аналоговое качество передачи образной картинки, которое добивались талантливые художники с помощью кисти и красок с давних времён. Но когда появляется (XVI век?) иллюстрированный кодекс, написанный неизвестным автором на неизвестном языке с использованием неизвестного алфавита (Манускрипт Войнича), становится понятным как достигается картинкой с текстом бесконечность значений меры образа. Если некоторые скептики до сих пор рассматривают рукопись Войнича как мистификацию, то Луиджи Серафини недавно создал энциклопедию воображаемого мира Codex Seraphinianus (1981) в 4К по примеру средневековых научных кодексов: каждая страница детально изображает определенный объект, действие или явление; аннотации написаны на выдуманном языке. Следовательно, существование бесконечнозначных и бесконечномерных образных методов измерения качественных значений нечисловых переменных это реальность. Этим методом живая клетка создаёт образы самой себя, других клеток, тканей и органов. Это происходит с помощью везикул, которые любая живая клетка производит и поглощает постоянно. Конечно же, сам "язык", химические краски и биологический образ этой картины понятны только самой живой клетке. Именно так и пишется родословная каждой клеткой в собственной клеточной книге (CellBook), которая состоит из генома («органа клетки» по определению Барбары МакКлинток), цитоплазмы клетки со всем молекулярным содержимым и живой мембраны (Senome=Сеном), которая своими мерцаниями помогает везикулам "рисовать" картину жизни. Peter Belobrov 18 Nov 2022 10:34 PIT00517 Интеграция данных из мультиомики Интеграция данных (образных биологических мер) из мультиомики - так можно определить второй день вебинара.  Data integration from multiomics to imaging | Day 2/2 | 2022 Graham Johnson, Allen Institute for Cell Science, Welcome and introductory remarks 2022 Seattle Cell Science Symposium - Day 2, t=3.18-21.06 смотреть и слушать надо очень внимательно, т.к. Грэм Джонсон (Graham Johnson) показал 81 слайд за 18 минут, делая вводный доклад [1] {Pollard2017} Thomas D. Pollard, William C. Earnshaw, Jennifer Lippincott-Schwartz, and Graham Johnson. Cell Biology, 3rd Ed. Elsevier, 2017. 900 p. Peter Belobrov 31 Jan 2022 06:32 Brian Beliveau, University of Washington 2022 Seattle Cell Science Symposium - Day 1, t=1.06.38-1.26.16 “Oligo-based technologies for programmed spatial biology” = «Олиготехнологии для программируемой пространственной биологии». Аннотация: Такие подходы, как геномика, транскриптомика и протеомика, могут предоставить обширную информацию о наличии и изобилии биомолекул в больших популяциях клеток, а в последнее время даже в отдельных клетках. Однако как ансамблевые, так и одноклеточные версии этих методов требуют диссоциации сложных структур, таких как ткани, во время их экспериментальных рабочих процессов, что приводит к потере пространственной информации. Мультиплексные подходы к визуализации, способные визуализировать несколько видов ДНК, РНК или белков в одном и том же образце, могут обеспечить ценный дополнительный подход к методам «омики», особенно в контексте тканей. Мы представили SABRE — усиление сигнала за счет реакции обмена. SABER обеспечивает мультиплексную амплификацию ДНК и РНК флуоресцентной гибридизации in situ (FISH) и сигналов иммунофлуоресценции в фиксированных клетках и тканях, позволяя картировать пространственные паттерны экспрессии генов и белков и организацию хромосом в их естественном контексте. Мы также представили «PaintSHOP», интерактивный веб-ресурс, который облегчает разработку и заказ наборов зондов FISH на основе олигонуклеотидов. Вместе эти инструменты предоставляют исследователям доступную и модульную основу для проведения экспериментов по созданию пространственных изображений. Библиография [1] {Hershberg2021} Hershberg, Elliot A., Conor K. Camplisson, Jennie L. Close, Sahar Attar, Ryan Chern, Yuzhen Liu, Shreeram Akilesh, Philip R. Nicovich, and Brian J. Beliveau. PaintSHOP enables the interactive design of transcriptome-and genome-scale oligonucleotide FISH experiments // Nature methods 18 (8), 937-944 (2021). doi 10.1038/s41592-021-01187-3 [2] {Martin2022v4} Martin, Beth K., Chengxiang Qiu, Eva Nichols, Melissa Phung, Rula Green-Gladden, Sanjay Srivatsan, Ronnie Blecher-Gonen et al. An optimized protocol for single cell transcriptional profiling by combinatorial indexing // Preprint arXiv 2110.15400 (2021). [Submitted on 28 Oct 2021 (v1), last revised 6 Jan 2022 (this version, v4)] Peter Belobrov 31 Jan 2022 06:08 PIT00515 Митофагия в первичных нейронах Chantell Evans, Duke University 2022 Seattle Cell Science Symposium - Day 1, t=46.00-1.06.38 “Investigating the Spatiotemporal dynamics of neuronal mitophagy” = «Исследование пространственно-временной динамики нейрональной митофагии». Доклад основан на статьях [6, 7] и более поздних работах Шантелл Эванс. Аннотация: Поврежденные митохондрии удаляются из клетки посредством митофагии. Этот путь может быть важен для гомеостаза нейронов, поскольку мутации в компонентах пути вызывают болезнь Паркинсона (БП) и боковой амиотрофический склероз (БАС). Мы использовали визуализацию в реальном времени для исследования пространственно-временной динамики митофагии в первичных нейронах после легкого окислительного стресса. Белки, ассоциированные с митофагией, быстро транслоцируются в деполяризованные митохондрии, и митохондрии изолируются в аутофагосомах в течение часа после повреждения. Удивительно, но нижестоящая деградация поглощенных митохондрий была отсрочена, прежде всего, из-за медленного подкисления образовавшихся митофагосом. Экспрессия мутации, связанной с БАС, нарушала функцию митохондриальной сети, а стресс усугублял этот эффект. Эти результаты свидетельствуют о том, что медленное обновление поврежденных митохондрий может повышать восприимчивость нейронов к нейродегенерации. Amyotrophic lateral sclerosis (ALS) = Боковой амиотрофический склероз (БАС) — изнурительное и неизлечимое заболевание, сопровождающееся потерей моторных нейронов и последующей атрофией мышц. Митофагия — избирательная деградация митохондрий путем аутофагии. Это часто происходит с дефектными митохондриями после повреждения или стресса. Процесс митофагии был впервые описан более ста лет назад Маргарет Рид Льюис и Уоррен Хармон Льюис [1]. Эшфорд и Портер использовали электронную микроскопию для наблюдения фрагментов митохондрий в лизосомах печени к 1962 г. [2] и в 1977 г. было высказано предположение, что «в митохондриях развиваются функциональные изменения, которые активируют аутофагию» [3]. Термин «митофагия» используется с 1998 г. [4]. Митофагия играет ключевую роль в поддержании здоровья клетки. Он способствует обороту митохондрий и предотвращает накопление дисфункциональных митохондрий, которые могут привести к клеточной дегенерации. Он опосредуется Atg32 (у дрожжей) и NIX и его регулятором BNIP3 у млекопитающих. Митофагия регулируется белками PINK1 и паркином. В дополнение к избирательному удалению поврежденных митохондрий митофагия также необходима для приспособления количества митохондрий к меняющимся метаболическим потребностям клеток, для стабильного митохондриального обмена и на определенных стадиях клеточного развития, например, во время клеточной дифференцировки эритроцитов [5]. Библиография [1] {Lewis1915} Lewis, Margaret Reed, and Warren Harmon Lewis. Mitochondria (and other cytoplasmic structures) in tissue cultures // American Journal of Anatomy 17 (3), 339-401 (1915). doi 10.1002/aja.1000170304. Cited by 340 [2] {Ashford1962} Ashford, Thomas P., and Keith R. Porter. Cytoplasmic components in hepatic cell lysosomes // The Journal of cell biology 12 (1), 198-202 (1962). doi 10.1083/jcb.12.1.198. Cited by 1077 [3] {Beaulaton1977} Beaulaton, Jacques, and Richard A. Lockshin. Ultrastructural study of the normal degeneration of the intersegmental muscles of Antheraea polyphemus and Manduca sexta (Insecta, Lepidoptera) with particular reference to cellular autophagy // Journal of morphology 154 (1), 39-57 (1977). doi 10.1002/jmor.1051540104. Cited by 155 [4] {Scott1988} Scott, Sidney V., and Daniel J. Klionsky. Delivery of proteins and organelles to the vacuole from the cytoplasm // Current opinion in cell biology 10 (4), 523-529 (1998). doi 10.1016/S0955-0674(98)80068-9. Cited by 105 [5] {Youle2011} Youle, Richard J., and Derek P. Narendra. Mechanisms of mitophagy // Nature reviews Molecular cell biology 12 (1), 9-14 (2011). doi 10.1038/nrm3028. Cited by 2699 [6] {Evans2019} Evans, Chantell S., and Erika LF Holzbaur. Autophagy and mitophagy in ALS // Neurobiology of disease 122, 35-40 (2019). doi 10.1016/j.nbd.2018.07.005 Cited by 82 Аннотация: Боковой амиотрофический склероз (БАС) — изнурительное и неизлечимое заболевание, сопровождающееся потерей моторных нейронов и последующей атрофией мышц. Генетические исследования выявили дефицит аутофагии и/или митофагии в начале заболевания. Здесь мы рассмотрим недавний прогресс в нашем понимании путей аутофагии и митофагии в нейронах и того, как на эти пути могут влиять мутации в генах, включая DCTN1, OPTN, TBK1, VCP и C9ORF72. Мы также обсуждаем последствия модуляции аутофагии при БАС, подчеркивая как потенциал этого подхода, так и опасения, возникающие при нацеливании на этот путь в качестве терапевтической стратегии при нейродегенеративных заболеваниях. [7] {Evans2020} Review Evans, Chantell S., and Erika LF Holzbaur. Quality control in neurons: mitophagy and other selective autophagy mechanisms // Journal of molecular biology 432 (1), 240-260 (2020). doi 10.1016/j.jmb.2019.06.031 Cited by 37 Аннотация: Специфическое удаление органелл посредством селективной аутофагии важно для поддержания гомеостаза нейронов. Генетические исследования показывают, что дефицит этих путей связан с нейродегенеративными заболеваниями, включая болезнь Паркинсона и боковой амиотрофический склероз. Здесь мы рассмотрим наше текущее понимание путей, которые регулируют контроль качества митохондрий, и сравним эти механизмы с механизмами, регулирующими оборот эндоплазматического ретикулума и клиренс белковых агрегатов. Исследования показывают, что существует множество механизмов, регулирующих деградацию определенных частиц клетки, таких как дисфункциональные органеллы и белковые агрегаты. Эти механизмы имеют решающее значение для здоровья нейронов, поскольку нейроны однозначно уязвимы к нарушению путей контроля качества органелл из-за их морфологии, размера, полярности и постмитотической природы. Мы подчеркиваем последствия нарушения регуляции селективной аутофагии в нейронах и обсуждаем текущие проблемы, связанные с корреляцией неконгруэнтных результатов, полученных в системах in vitro и in vivo. 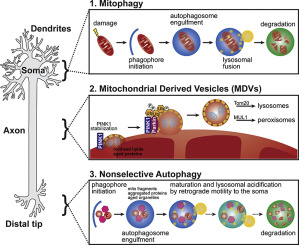 Graphical Abstract Peter Belobrov 31 Jan 2022 03:49 PIT00514 Чем определяется состояние клетки? «Клеточное состояние определяет мультимодальный сигнальный ответ одиночных клеток» - написали Бернхард Крамер и Лукас Пелкманс в 2019 [1]. Препринт цитировали 3 раза. Журнальный вариант читали пока только рецензенты. Доклад Лукаса не случайно оргкомитет поставил первым. Впрочем, давайте без суеты: посмотрим доклад, послушаем, подумаем, почитаем препринт ... Lucas Pelkmans, University of Zurich 2022 Seattle Cell Science Symposium - Day 1, t=15.20-45.48 “Multimodal perception links cellular state to decision making in single cells” = «Мультимодальное восприятие связывает клеточное состояние с принятием решений в отдельных клетках». Аннотация: Фундаментальное свойство клеток заключается в том, что они принимают решения, адаптированные к своему внутреннему состоянию и окружающей среде. Это контекстно-зависимое поведение требует обработки больших объемов информации, но неясно, как клетки могут надежно достичь этого, используя гетерогенные ответные сигналы. Чтобы изучить способность эпителиальных клеток человека обрабатывать информацию, мы здесь применяем стимуляцию эпидермального фактора роста в сочетании с мультиплексной количественной оценкой сигнальных ответов и несколькими маркерами клеточного состояния в нескольких пространственных масштабах. Мы обнаружили, что сигнальные узлы в сети отображают адаптивную обработку информации, что приводит к разнородным ответам факторов роста и позволяет узлам собирать частично неизбыточную информацию о состоянии сотовой связи. В совокупности, как мультимодальное восприятие, это предоставляет отдельным клеткам большой объем информации, чтобы точно определить концентрацию фактора роста в контексте их клеточного состояния и принимать решения, зависящие от клеточного состояния. Мы предполагаем, что неоднородность и сложность сигнальных сетей сопутствовали друг другу, чтобы обеспечить возможность принятия специфических и контекстно-зависимых сотовых решений в многоклеточных условиях. Библиография [1] {Kramer2019} Kramer, Bernhard A., and Lucas Pelkmans. Cellular state determines the multimodal signaling response of single cells // Preprint bioRxiv (2019). 55 p. doi 10.1101/2019.12.18.880930 Cited by 3 Аннотация: Многочисленные фундаментальные биологические процессы требуют, чтобы отдельные клетки правильно интерпретировали поступающие сигналы и точно реагировали на них. Как внутриклеточные сигнальные сети достигают интеграции сложной информации из различных контекстов, остается неясным. Здесь мы количественно оцениваем индуцированную эпидермальным фактором роста (EGF) гетерогенную активацию множества сигнальных белков, а также маркеров клеточного состояния в одних и тех же отдельных клетках в разных пространственных масштабах. Мы обнаружили, что острая реакция каждого узла в сигнальной сети тесно связана с клеточным состоянием частично неизбыточным образом. Это создает мультимодальный ответ, который лучше воспринимает разнообразие клеточных состояний, чем любой индивидуальный ответ, и позволяет отдельным клеткам точно определять концентрацию фактора роста в контексте их клеточного состояния. Обсуждение: Мы здесь количественно, in situ = на месте и во многих пространственных масштабах, резкие изменения в активированном состоянии 7 киназ, 2 транскрипционных факторов и 1 рибосомной субъединицы при воздействии EGF, а также 18 неизменяющихся маркеров цитоскелета, мембранного переноса, процессинга цитоплазматической РНК и механического, пролиферативное, метаболическое и транскрипционное состояние одновременно в отдельных клетках. Полученный набор данных показывает сильную связь между клеточным состоянием и реакцией отдельных клеток на входящий сигнал. Этот эффект возникает из-за нескольких пространственных масштабов и в совокупности объясняет большую часть гетерогенности, наблюдаемой в ответах одиночных клеток на воздействие фактора роста. Это показывает, что сигнальные узлы в сети не одинаково связаны с одними и теми же клеточными свойствами. и что изменчивость выходного отклика на одинаковый вход, отображаемый отдельными узлами, кодирует частично неизбыточную информацию. В совокупности неизбыточная информация, содержащаяся в мультимодальном ответе, является более точным представлением клеточного состояния, чем каждый отдельный ответ по отдельности. Кроме того, отдельные узлы демонстрируют диапазоны чувствительности, зависящие от клеточного состояния, при различных входных концентрациях фактора роста. Это говорит о том, что мультимодальный, зависящий от клеточного состояния характер передачи сигналов в организмах млекопитающих развился, чтобы наделить отдельные клетки высокой способностью к обработке информации, чтобы точно интерпретировать широкий спектр входной информации в сложном ландшафте клеточных состояний и принимать соответствующие меры (и придумывать правильное) решение. Интересно, что обнаруженные здесь свойства внутриклеточной обработки сигналов аналогичны мультимодальному восприятию, осуществляемому сенсорными системами в царстве многоклеточной жизни [31,, 53-56]. Оба отображают восприятие через несколько модальностей (множество сигнальных узлов по сравнению с несколькими сенсорными нейронами) с частичной неизбыточностью, что позволяет интегрировать несколько источников информации в более полную картину реальности. ключевые слова для понимания этой статьи: Peter Belobrov 30 Jan 2022 13:59 PIT00512 Анализ изображений ядра в hiPSC 10:35-11:05am) - Nuclear Morphogenesis Team, Allen Institute for Cell Science 2022 Seattle Cell Science Symposium - Day 2, 2.21.40-2.50.00 “Exploring mechanisms of nuclear shape and growth dynamics in hiPSC via integrated image analysis” = «Изучение механизмов формирования и динамики роста ядра в hiPSC с помощью интегрированного анализа изображений». Peter Belobrov 29 Jan 2022 23:44 PIT00511 Адаптация актиновыми сетями 10:15-10:35am) - Matthew Akamatsu, University of Washington. 2022 Seattle Cell Science Symposium - Day 2, 1.57.24-2.20.34 "Self-organization and load adaptation by endocytic actin networks" «Самоорганизация и адаптация к нагрузке эндоцитарными актиновыми сетями». Аннотация: ансамбли полимеризующихся белковых комплексов цитоскелета собираются в клетках, создавая направленную силу на клеточные мембраны. Во время клатрин-опосредованного эндоцитоза сети разветвленных актиновых филаментов собираются на плазматической мембране, чтобы толкать, тянуть и изменять форму эндоцитарной ямки. Лежащие в основе принципы, которые обеспечивают самосборку поляризованных разветвленных актиновых сетей в стохастической клеточной среде, не совсем понятны. Интеграция количественных моделей с количественными экспериментами позволяет нам итеративно предлагать и проверять такие механистические гипотезы. Мы объединили компьютерное моделирование на основе агентов с количественной флуоресцентной микроскопией живых клеток индуцированных плюрипотентных стволовых клеток человека с отредактированным геномом, чтобы определить механизмы, с помощью которых разветвленные актиновые сети организуются для создания силы в местах эндоцитоза млекопитающих. Мы ограничили нашу модель измерениями очищенных белков и проверили механистические предсказания с помощью микроскопии сверхвысокого разрешения и криоэлектронной томографии. С помощью этой комбинации подходов мы обнаружили, что расположение ключевых актин-связывающих белков в местах эндоцитоза позволяет разветвленным актиновым сетям самоорганизовываться в поляризованный массив, радиально сфокусированный на покрытой клатрином ямке. Такая организация филаментов позволяет сети динамически реагировать на увеличение нагрузки, например, на повышенное натяжение мембраны, за счет зародышеобразования дополнительных филаментов и накопления упругой энергии в согнутых филаментах. Регулярное связывание актин-ветвящихся и связывающих белков на актиновых филаментах ведет к разреженным асимметричным актиновым сетям, которые за счет постоянного повторного связывания с ямкой и перестройки производят достаточную силу для эндоцитарной интернализации. Peter Belobrov 29 Jan 2022 23:34 PIT00510 Созидание цитоскелета клетки 9:55-10:15am - Sabine Petry, Princeton University. 2022 Seattle Cell Science Symposium - Day 2, 1.30.14-1.56.44 "How to make microtubules and build the cytoskeleton" = «Как создаются микротрубочки и строится цитоскелет». Аннотация: Как клетка строит свой цитоскелет из микротрубочек? В соответствии с принципом Фейнмана «то, что я не могу создать, я не понимаю», моя лаборатория занимается этим вопросом, создавая механизм сегрегации хромосом с нуля. Сначала я расскажу вам, как формируется каркас микротрубочек в клетке. Расшифровав функцию наиболее важных вспомогательных белков микротрубочек, я представлю, как мы используем эти строительные блоки для воссоздания субструктуры веретена in vitro и определения плана ее построения. Наконец, я объясню, как мы комбинируем подструктуры веретена, как кусочки головоломки, чтобы собрать и, таким образом, понять функционирующее веретено, которое разделяет хромосомы. Изучая, как устроен цитоскелет МТ, я надеюсь помочь объяснить, как сотни белков могут самостоятельно собираться в нанометровом масштабе в сложную молекулярную машину, в 1000 раз превышающую размеры ее составляющих, что является проблемой для биохимии 21 века. Peter Belobrov 29 Jan 2022 23:24 PIT00509 Клетки в пространстве органа 9:25-9:45am) - Shila Ghazanfar, University of Cambridge. 2022 Seattle Cell Science Symposium - Day 2, 1.03.16-1.22.22 “Data integration for molecule-resolved spatial gene expression of mouse organogenesis” = «Интеграция данных для экспрессии пространственного гена с молекулярным разрешением в органогенезе мыши». Аннотация: Профилирование диссоциированной транскрипции одиночных клеток расширило наши знания о молекулярных основах развития. Новые технологии пространственной транскриптомики, такие как seqFISH, позволяют точно обнаруживать молекулы РНК в их естественном пространственном контексте. Здесь мы применили seqFISH для создания пространственной карты органогенеза мыши с высоким разрешением, с сотнями генов-мишеней, захваченных в срезах тканей. Путем компьютерной интеграции этого пространственного контекста и мультиплексных измерений транскрипции с транскриптомными атласами отдельных клеток в целом мы можем выявить направления клеточной дифференцировки, которые не сразу очевидны из любой технологии в отдельности. 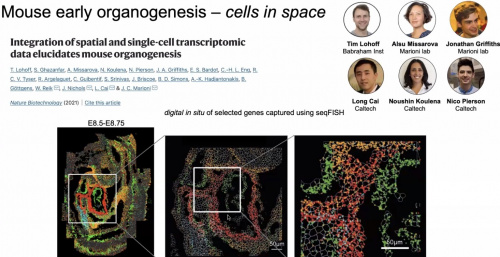 Ранний органогенез мыши - клетки в пространстве органа Библиография {Lohoff2022} Lohoff, T., S. Ghazanfar, A. Missarova, N. Koulena, N. Pierson, J. A. Griffiths, E. S. Bardot et al. Integration of spatial and single-cell transcriptomic data elucidates mouse organogenesis // Nature biotechnology 40 (1), 74-85 (2022). doi 10.1038/s41587-021-01006-2 Peter Belobrov 29 Jan 2022 22:56 PIT00508 Пространственная мультиомика 9:05-9:25am) - Sanja Vickovic, Broad Institute. 2022 Seattle Cell Science Symposium - Day 2, 43.18-1.02.20 “SM-Omics as an automated platform for high-throughput spatial multi-omics” = «SM-Omics как автоматизированная платформа для высокопроизводительной пространственной мультиомики». Аннотация: Пространственная организация клеток и молекул играет ключевую роль в функционировании тканей при гомеостазе и заболеваниях. Пространственная транскриптомика недавно стала ключевой техникой для захвата и позиционного штрих-кодирования РНК непосредственно в тканях. Мы продвигаем применение пространственной транскриптомики в масштабе, представляя Spatial Multi-Omics (SM-Omics) как полностью автоматизированную, высокопроизводительную платформу на основе секвенирования для комбинированных и пространственно разрешенных транскриптомных измерений и измерений белков на основе антител. SM-Omics использует антитела со штрих-кодом ДНК, иммунофлуоресценцию или их комбинацию для масштабирования и объединения пространственной транскриптомики и пространственного мультиплексного обнаружения белков на основе антител. SM-Omics позволяет обрабатывать до 64 пространственных реакций in situ или до 96 готовых к секвенированию библиотек высокой сложности в течение примерно 2 дней. Peter Belobrov 29 Jan 2022 22:02 PIT00507 3D-модели целой бактериальной клетки Это пост посвящен Дэвиду Гудселлу (David Goodsell). 8:45-9:05am - David Goodsell, Scripps Research. 2022 Seattle Cell Science Symposium - Day 2, 21.08-43.02 “Building Structural Models of a Whole Bacterial Cell” = «Построение структурных моделей целой бактериальной клетки». Аннотация: Основываясь на предыдущих художественных подходах к визуализации молекулярной структуры живых клеток, мы создали 3D-модели целой бактериальной клетки, включающей все макромолекулы. Концентрация, положения и взаимодействия молекул взяты из последних компьютерных моделей биологии целых клеточных систем, интегрированы с экспериментальными и смоделированными по гомологии структурами 482 белковых мономеров и 201 белковых комплексов, а также основанными на решетке моделями генома, РНК и связанных с ними структур. макромолекулы. Был разработан вычислительный рабочий процесс для упрощения сбора и курирования этих структур, определения «рецепта», определяющего состав клеточной модели, а также построения и оптимизации моделей на основе этого рецепта. Также были разработаны методы визуализации, позволяющие исследовать функциональные характеристики генома и протеома. См. сайт Гудселла: Welcome to my new website!) To reflect the expanding interests of our labs, Art Olson, Stefano Forli, Michel Sanner and I have recently rebranded the Molecular Graphics Laboratory as the Center for Computational Structural Biology (CCSB). Основные книги Гудселла: David S. Goodsell. Bionanotechnology: Lessons from Nature. Wiley-Liss, 2004. 347 p. David S. Goodsell. The Machinery of Life. 2nd Ed. Copernicus Books, 2009. 170 p. David S. Goodsell. Atomic Evidence: Seeing the Molecular Basis of Life. Copernicus, Year: 2016. 180 p. Peter Belobrov 29 Jan 2022 21:34 PIT00506 Протеомика биологических переменных В единственном докладе было использовано моё любимое понятие "биологическая переменная" на примере использования протеомики для анализа возраста как биологической переменной. 10:45-11:05am - Devin Schweppe, University of Washington 2022 Seattle Cell Science Symposium - Day 1, 2.21.25-2.41.48 “Multiplexed Mapping of Complex Proteomes” = «Мультиплексное картирование сложных протеомов» Peter Belobrov 29 Jan 2022 21:14 PIT00505 4D CELL Biology Lab  Результаты и перспективы программы исследований (PI) Йоханнеса Шёнеберга (Johannes Schöeneberg) были представлены в докладе Johannes Schöeneberg, University of California San Diego. “Integration of particle simulations, lattice light-sheet imaging and deep learning in quantitative cell biology” = «Интеграция моделирования частиц, визуализации решетчатых осветлённых листов изображений и глубокого обучения в количественной клеточной биологии». 2022 Seattle Cell Science Symposium - Day 1, t=1.59.30-2.20.50 Peter Belobrov 29 Jan 2022 17:58 PIT00504 Эпителиально-мезенхимальный переход Лучшая картинка для прояснения сути дела (англ. Wiki - перевода пока нет): 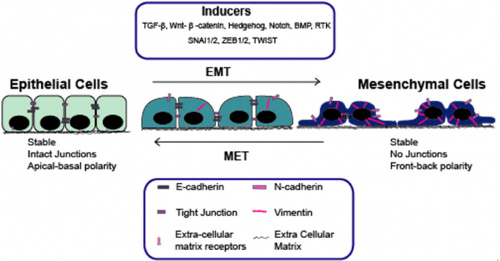 EMT - epithelial-mesenchymal transition; MET - mesenchymal-epithelial transition 9:55-10:25am - Epithelial to mesenchymal transition (EMT) Team Talk, Allen Institute for Cell Science 2022 Seattle Cell Science Symposium - Day 1, 1.32.00-1.59.28 “Using the epithelial to mesenchymal transition (EMT) in human induced pluripotent stem cells (hiPSC) as a model to study cell states and state transitions by combining cell behavior, cell organization, and cell identity” = «Использование эпителиально-мезенхимального перехода (EMT) в индуцированных плюрипотентных стволовых клетках (hiPSC) человека в качестве модели для изучения клеточных и переходных состояний клеток путем объединения поведения клеток, клеточной организации и клеточной идентичности». Эпителиально-мезенхимальный переход (ЕМТ). Внимание: 1320 статей в 2022 на англ., конечно, в Google Scholar. На русском 269 статей всего! Дворяшина, И.А., Ю.И. Великородная, А.В. Терентьев, В.Л. Загребин. ЭПИТЕЛИАЛЬНО-МЕЗЕНХИМАЛЬНЫЙ ПЕРЕХОД I ТИПА КАК ВАЖНЫЙ БИОЛОГИЧЕСКИЙ ПРОЦЕСС В ЭМБРИОГЕНЕЗЕ // Вестник Волгоградского гос.мед.ун-та 2 (78), 37-45 (2021). Аннотация. Эпителиально-мезенхимальный (ЭМП) и мезенхимально-эпителиальный (МЭП) переходы играют значительную роль в процессах эмбрио-, органогенеза и клеточной дифференцировке. Эпителиально-мезенхимальный переход также способствует регенерации поврежденных тканей, но в случае его аберрантной активации может инициировать и усугублять течение таких патологических процессов, как фиброз, метастазирование и канцерогенез. В обзоре мы рассмотрели основные исторические этапы изучения данных процессов в контексте эмбрионального развития, а также обсудили проблемы терминологии и разные взгляды на представления о детерминированности зрелых клеток. Ключевые слова: эпителиально-мезенхимальный переход, эмбриогенез, мезенхима, эктодерма. Peter Belobrov 28 Jan 2022 13:42 PIT00502 Пространственная биология клетки Паттерны пространственной биологии клетки в проекте родословной после конференции надо существенно добавить новыми методами и образными биологическими мерами. Peter Belobrov 13 Jan 2022 12:44 Seattle Cell Science Symposium 27-28/1/2022 прошел, «и вновь я посетил», и за две ночи наполнился идеями Института клеточных наук Аллена, который показал высший класс науки о клетках, и обновил программой этот пост. 1-ый день вебинара 2022 Seattle Cell Science Symposium - Day 1 в Сиэтле (27/1/2022) прошел по следующей программе: 8:30am - Welcome, Ru Gunawardane, Executive Director, Allen Institute for Cell Science [Day1, t=4:50] 8:30-8:45am - Susanne Rafelski, Allen Institute for Cell Science, Introductory remarks см PIT00514 пост (8:45-9:05am) - Lucas Pelkmans, University of Zurich 2022 Seattle Cell Science Symposium - Day 1, t=15.20-45.48 “Multimodal perception links cellular state to decision making in single cells” = «Мультимодальное восприятие связывает клеточное состояние с принятием решений в отдельных клетках». см PIT00515 пост (9:05-9:25am) - Chantell Evans, Duke University 2022 Seattle Cell Science Symposium - Day 1, t=46.00-1.06.38 “Investigating the Spatiotemporal dynamics of neuronal mitophagy” = «Исследование пространственно-временной динамики нейрональной митофагии». см PIT00516 пост (9:25-9:45am) - Brian Beliveau, University of Washington 2022 Seattle Cell Science Symposium - Day 1, t=1.06.38-1.26.16 “Oligo-based technologies for programmed spatial biology” = «Олиготехнологии для программируемой пространственной биологии». 9:45-9:55am - Break см PIT00504 пост (9:55-10:25am) - Epithelial to mesenchymal transition (EMT) Team Talk, Allen Institute for Cell Science 2022 Seattle Cell Science Symposium - Day 1, t=1.32.00-1.59.28 “Using the epithelial to mesenchymal transition (EMT) in human induced pluripotent stem cells (hiPSC) as a model to study cell states and state transitions by combining cell behavior, cell organization, and cell identity” = «Использование эпителиально-мезенхимального перехода (EMT) в индуцированных плюрипотентных стволовых клетках (hiPSC) человека в качестве модели для изучения клеточных и переходных состояний клеток путем объединения поведения клеток, клеточной организации и клеточной идентичности». см PIT00505 пост (10:25-10:45am) - Johannes Schöeneberg, University of California San Diego. 2022 Seattle Cell Science Symposium - Day 1, t=1.59.30-2.20.50 “Integration of particle simulations, lattice light-sheet imaging and deep learning in quantitative cell biology” = «Интеграция моделирования частиц, визуализации решетчатых осветлённых листов изображений и глубокого обучения в количественной клеточной биологии» см PIT00506 пост (10:45-11:05am) - Devin Schweppe, University of Washington 2022 Seattle Cell Science Symposium - Day 1, t=2.21.25-2.41.48 “Multiplexed Mapping of Complex Proteomes” = «Мультиплексное картирование сложных протеомов» 11:05-11:45am - Panel Discussion 11:45-11:50am - Closing Remarks 2-ый день вебинара 2022 Seattle Cell Science Symposium - Day 2 по клеточным наукам симпозиума в Сиэтле (27/1/2022) прошел по следующей программе: Сегодня программа ещё более интересная и такая: 8:25-8:30am - Virtual meeting log-in см PIT00517 пост (8:30-8:45am) - Graham Johnson, Allen Institute for Cell Science, Welcome and introductory remarks 2022 Seattle Cell Science Symposium - Day 2, t=3.18-21.06 см PIT00507 пост (8:45-9:05am) - David Goodsell, Scripps Research. 2022 Seattle Cell Science Symposium - Day 2, t=21.08-43.02 “Building Structural Models of a Whole Bacterial Cell” = «Построение структурных моделей целой бактериальной клетки» см PIT00508 пост (9:05-9:25am) - Sanja Vickovic, Broad Institute. 2022 Seattle Cell Science Symposium - Day 2, t=43.18-1.02.20 “SM-Omics as an automated platform for high-throughput spatial multi-omics” = «SM-Omics как автоматизированная платформа для высокопроизводительной пространственной мультиомики». см PIT00509 пост (9:25-9:45am) - Shila Ghazanfar, University of Cambridge. 2022 Seattle Cell Science Symposium - Day 2, t=1.03.16-1.22.22 “Data integration for molecule-resolved spatial gene expression of mouse organogenesis” = «Интеграция данных для экспрессии пространственного гена с молекулярным разрешением в органогенезе мыши». см PIT00510 пост (9:55-10:15am) - Sabine Petry, Princeton University. 2022 Seattle Cell Science Symposium - Day 2, t=1.30.14-1.56.44 "How to make microtubules and build the cytoskeleton" «Как сделать микротрубочки и построить цитоскелет». см PIT00511 пост (10:15-10:35am) - Matthew Akamatsu, University of Washington. 2022 Seattle Cell Science Symposium - Day 2, t=1.57.24-2.20.34 "Self-organization and load adaptation by endocytic actin networks" «Самоорганизация и адаптация к нагрузке эндоцитарными актиновыми сетями». см PIT00512 пост (10:35-11:05am) - Nuclear Morphogenesis Team, Allen Institute for Cell Science 2022 Seattle Cell Science Symposium - Day 2, t=2.21.40-2.50.00 “Exploring mechanisms of nuclear shape and growth dynamics in hiPSC via integrated image analysis” = «Изучение механизмов формирования и динамики роста ядра в hiPSC с помощью интегрированного анализа изображений» 11:05-11:45am - Panel Discussion 11:45-11:50am - Closing Remarks Peter Belobrov 14 Dec 2021 15:38 PIT00494 Сложное действие Чтобы как-то продвинуться в теории сложных взаимодействий (пост PIT00168 от 21/12/2014), надо разработать теорию сложного действия. Эта мысль появлялась много раз, хотя все основные соображения локализовались вокруг бесконечномерных и бесконечнозначных образных переменных. Peter Belobrov 31 Oct 2020 14:50 PIT00482 Тонкости «созидающих» кодов AAR в канале семинаров высказал интересные мысли. Как описывать поведение (алгоритмы) без использования языка программирования? Хороший перевод доклада Лесли Лэмпорта позволил «созидающие коды» высветить ещё ярче так. Существует принципиальная разница в описании поведения «до того» и «после того». Анекдотический ответ «вместо того» создаёт правило точнейшего понимания тонкостей отличия алгоритма от описания творческого или созидательного поведения. Предполагаю, что именно здесь DIA найдёт подсказку для «созидающих кодов». По поводу вычислений на квантовом компьютере Google осталась информация здесь: [Arute2019], здесь: О «квантовом превосходстве» и здесь: Квантовое превосходство наступает, но это не изменит мир. Думаю, что наш подход к биологическим измерениям качественных свойств живых систем правильный и может быть предварительно определён как превосходство аналоговых вычислений над квантовыми, если не беспокоиться «совершенно дурацким» шифрованием, понимая, что биологическая эволюция в собственных «созидающих кодах» шифрование не применяет. Однако химические [Barbieri2003] и биологические [Barbieri2015] коды используются для перезаписывания генома при каждом клеточном делении [Shapiro2013]. Ссылки {Arute2019} F. Arute, K. Arya, R. Babbush et al. Quantum supremacy using a programmable superconducting processor // Nature 574, 505–510 (2019). doi 10.1038/s41586-019-1666-5. {Barbieri2003} Barbieri M. The organic codes. An introduction to semantic biology. Cambridge, UK: Cambridge University Press. 2003. 317 p. {Barbieri2015} Marcello Barbieri. Code Biology: A New Science of Life. Springer International Publishing, 2015. 236 p. {Shapiro2013} James A. Shapiro. How life changes itself: The Read–Write (RW) genome // Physics of Life Reviews, 10 (3), 287–323 (2013). doi 10.1016/j.plrev.2013.07.001 PDF uchicago.edu Peter Belobrov 07 Dec 2019 14:06 PIT00473 Профили интронов и ... новый прорыв Пожалуйста, прежде, чем читать дальше, освежите в памяти понятия: сплайсинг. экзон и интрон. Новая методика определение последовательности интронов флуоресцентной гибридизацией in situ, названная intron seqFISH, позволяет по профилям интронов идентифицировать типы и состояния клеток. Профилирование около половины от общего числа генов у млекопитающих (мыши) позволяет отображать более 10 000 генов в отдельных клетках. Год назад мало кто поверил в этот поистине фантастический результат [Shah2018], который будет рассказан на 2019 Allen Frontiers Symposium in Boston 16-17/10/2019 подробно. Посмотрим на графическую аннотацию и резюме статьи [Shah2018]. стр 23 (продолжение последует вскоре) {Shah2018} Sheel Shah, Yodai Takei, Wen Zhou, Eric Lubeck, Jina Yun, Chee-Huat Linus Eng, Noushin Koulena et al. Dynamics and spatial genomics of the nascent transcriptome by intron seqFISH // Cell 174 (2), 363-376 (2018). doi 10.1016/j.cell.2018.05.035 Peter Belobrov 23 Aug 2019 12:16 PIT00471 Pro Git вам в помощь! МОЛПИТ -- единственная в мире команда, которая разрабатывает принципиально новый подход к будущей фундаментальной биологии. Если говорить кратко, то через родословную книги клетки. С точки зрения алгоритмов и программ в биологии Денис Нобл предложил ключевую идею, что традиционных компьютерных программ в живых системах нигде нет. Эта точка зрения критикуется, хотя нами уже сформулирована идея созидающих кодов и формировании собственных биологических алгоритмов в клетках, тканях, органах и организмах по мере биологического развития. Причём сами алгоритмы не наследуются, но наследуется родословная клеточных книг путём перемешивания. Как только это идея овладеет основными научными массами, развивающими фундаментальную биологическую науку, так сразу вспыхнут реальные биологические алгоритмы. Где и как это произойдёт - неизвестно, но это появится в ближайшее время. Можно даже сказать как. Понимание этого произойдёт на пути изучения транскриптомики одиночных клеток. Подробнее см посты PIT00470 и PIT00190. Скачайте себе книгу Pro Git by Scott Chacon and Ben Straub, 2nd Ed. на русском языке, сейчас версия 2.1.121-1-gabdb461, 27.01.2026. Два примера прояснят суть этого поста и смысл того, почему в происходящей биологической революции, причиной которой стало секвенирование РНК одиночных клеток (scRNA-seq), без Git-технологии, пожалуй, не разобраться. Dynamo: Mapping Vector Field of Single Cells Xiaojie Qiu, Yan Zhang, Dian Yang, Shayan Hosseinzadeh, Li Wang, Ruoshi Yuan, Song Xu et al. Mapping Vector Field of Single Cells // Preprint bioRxiv (2019): 696724. doi 10.1101/696724; Cited by 30 - цитирование прекратилось в 2024 году, так как отслеживание родословных в одноклеточной омикс-технологии интенсивно развивалось за последние пару лет PIT00649. Analysis of single cell RNA-seq data - двухдневный курс по анализу данных по секвенированию РНК одиночных клеток с двумя видео по 8 часов без понимания Git-технологии элементарно не освоишь. Действуйте. Рекомендуемая книга вам поможет. Мне она точно помогла вздохнуть свежим воздухом новой технологии обмена данными между разработчиками. Обновление 14/3/2026. Szałata, Artur, Karin Hrovatin, Sören Becker, Alejandro Tejada-Lapuerta, Haotian Cui, Bo Wang, and Fabian J. Theis. Transformers in single-cell omics: a review and new perspectives // Nature methods 21 (8), 1430-1443 (2024). хhttps://doi.org/10.1038/s41592-024-02353-z doi 10.1038/s41592-024-02353-z]; Cited by 158. Peter Belobrov 09 Aug 2019 21:04 PIT00470 Анализ РНК одной клетки Десятилетие секвенирование РНК одиночных клеток (scRNA-seq) с 2009 торжествует в науке, переделывая фундаментальную биологию по принципу тихой революции. Кто одноклеточным анализом РНК не владеет, тот работает в позапрошлом веке. А мы продолжаем выкладывать ссылки на лучшие источники, зная которые можно не отстать от развития современной биологии. Посмотрите пост PIT00190. Это одно из продолжений, которые следуют. Внимательный студент спросит в когда началось секвенирование ДНК одиночных клеток? Точного и простого ответа не будет. Причин много. Вот уже несколько месяцев я читаю самые новые статьи, которых всё больше и больше, которые всё глубже и глубже. Вот уже пришла необходимость самые новые публикации освещать в новых постах. См PIT00471 и далее ... Peter Belobrov 01 Aug 2019 01:44 Тип клетки – фундаментальная биологическая постоянная,принимающая конечное число нечисловых значений (в виде образов) и континуум физико-химических числовых значений.Рассмотрим Peter Belobrov 08 Jun 2019 19:08 Основная идея, которая поможет продвинуться в атласах клетокБудем различать биологические, химические, физические и математические переменные. Конечно, существует множество других переменных, как-то: лингвистические, семантические, философские и большое разнообразие различных величин, которые принимают нечисловые значения. Чаще всего это графические или смысловые образы, которые выражаются картинкой или ключевыми словами понятийного аппарата конкретных наук. Классификация по типам образных значений позволяет ввести нечисловые константы, например, тип клетки. Белая книга атласа клеток человека [Regev2018pr] Аннотация Атлас клеток человека (HCA = Human Cell Atlas) будет составлен из комплексных справочных карт всех клеток человека - основных единиц жизни - в качестве основы для понимания основных биологических процессов человека и диагностики, мониторинга и лечения заболеваний. Это поможет ученым понять, как генетические варианты влияют на риск заболевания, определить токсичность лекарств, найти лучшую терапию и продвинуть регенеративную медицину. Ресурс таких амбиций и масштабов должен создаваться поэтапно, увеличиваясь в размере, ширине и разрешении по мере развития технологий и углубления понимания. Поэтому мы будем проводить Фазу 1 как набор ведущих проектов в ключевых тканях, системах и органах. Мы соберем вместе экспертов в области биологии, медицины, геномики, разработки технологий и вычислений (включая анализ данных, разработку программного обеспечения и визуализацию). {Regev2018pr} Aviv Regev, Sarah Teichmann, Orit Rozenblatt-Rosen, Michael Stubbington, Kristin Ardlie, Ido Amit, Paola Arlotta et al. The Human Cell Atlas White Paper // Preprint arXiv:1810.05192 (v1 11 Oct 2018). Peter Belobrov 08 Jun 2019 18:30 PIT00456 Джеймс Шапиро: «Геном не остров» Ни один геном не является островом: на пути к пониманию эволюции в 21-м веке АннотацияОбычное эволюционное мышление 20-го века основывалось на идее изолированных геномов для каждого вида. Любая возможность вклада истории жизни в зародышевую линию была строго исключена доктриной Вейсмана, а изменение генома было приписано случайным ошибкам копирования. Сегодня мы знаем, что многие жизненные события приводят к быстрым и неслучайным эволюционным изменениям, обусловленным специфическими клеточными функциями. Существует множество способов взаимодействия геномов, вирусов, клеток и организмов для создания эволюционных изменений. К ним относятся слияния клеток и активация природной генной инженерии в результате стресса, инфекции и межвидовой гибридизации. Кроме того, мы знаем молекулярные механизмы передачи информации об истории жизни через поколения путём гамет. Эти открытия требуют новой программы эволюционной теории и новых экспериментальных проектов для изучения геномных воздействий стрессов, биотических взаимодействий и сенсорных воздействий, поступающих из окружающей среды. В обзоре предложены некоторые общие рекомендации по обогащению экспериментов по эволюции для включения новых знаний и поиска ответов на ранее не задаваемые вопросы.{Shapiro2019} James A. Shapiro. No genome is an island: toward a 21st century agenda for evolution // Annals of the New York Academy of Sciences 1447 (1), 21-52 (2019). doi 10.1111/nyas.14044. Peter Belobrov 29 May 2019 09:28 PIT00455 Мотивы факторов транскрипции Наконец-то, скажет любой нормальный биолог, иерархию и важность пространственных мотивов, в частности, для транскрипционных факторов удалось установить экспериментально. Анатолий Глянцев мгновенно сделал русский реферат статьи [Lambert2019], точнее пресс-релиза "установлен ряд генов, которые могут быть ключом к тому, как эволюционировали люди" и опубликовал неплохой перевод его Обнаружены десятки "менеджеров по генам", делающих нас людьми. Читайте и вникайте! Транскрипционные сигналы, факторы и сама регуляция транскрипции - важнейшие биологические процессы, понимание которых ещё недостаточно ясно охарактеризовано, хотя очень быстрый прогресс в исследовании деталей транскрипции. Так в недавнем обзоре "Геном не остров" Шапиро предложил много новых идей для понимания эволюции [Shapiro2019]. Этот обзор практически переведен. Сделана пара семинаров по этому обзору. {Lambert2019} Samuel A. Lambert, Ally W. H. Yang, Alexander Sasse, Gwendolyn Cowley, Mihai Albu, Mark X. Caddick, Quaid D. Morris, Matthew T. Weirauch & Timothy R. Hughes. Similarity regression predicts evolution of transcription factor sequence specificity // Nature Genetics 51, 981–989 (2019). doi 10.1038/s41588-019-0411-1 {Shapiro2019} James A. Shapiro. No genome is an island: toward a 21st century agenda for evolution // Annals of the New York Academy of Sciences (2019). ОБЗОР Первая публикация 22/3/2019 doi 10.1111/nyas.14044 Peter Belobrov 29 May 2019 01:30 PIT00451 Биология развития клеток Молекулярный контроль макроскопических сил способствует формированию органов позвоночных при эмбриональном развитии. Статья [Nerurkar2019] и пресс-релиз [Evarts2019] представили очень интересные результаты для размышлений. Молекулярные градиенты были установлены в качестве ключевой движущей силы клеточной организации в формировании ранних структур во время эмбрионального развития [Free2019].  Отслеживание движений клеток энтодермы из интервальных экспериментов, изучающих, как клетки становятся тканью (интернализуются) для формирования задней кишки у развивающегося эмбриона цыпленка; время кодируется цветом трека от 0 часов (фиолетовый) до 16 часов (белый) [Evarts2019]. 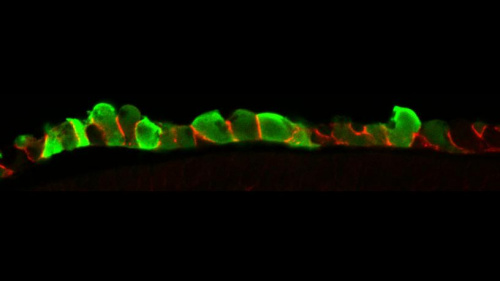 Микрофотография клеток энтодермы у эмбрионов кур. Клетки визуализируются путем направленной экспрессии зеленого флуоресцентного белка (зеленого); белок клеточной адгезии Е-кадгерин визуализируется красным цветом [Evarts2019]. (Для справки) FGF или Факторы роста фибробластов, относятся к семейству факторов роста, участвующих в ангиогенезе, заживлении ран и эмбриональном развитии. Факторы роста фибробластов — это гепарин-связывающие белки. Было доказано, что взаимодействия с расположенными на поверхности клеток протеогликанами необходимы для передачи сигнала факторов роста фибробластов. Факторы роста фибробластов играют ключевую роль в процессах пролиферации и дифференцировки широкого спектра клеток и тканей. Рекомендую пройти по ссылке SuppL Info и посмотреть 7 видео, чтобы у Вас сложился точный образ формирования ткани из клеток [Nerurkar2019]. Литература {Nerurkar2019} Nandan L. Nerurkar, ChangHee Lee, L. Mahadevan, and Clifford J. Tabin. Molecular control of macroscopic forces drives formation of the vertebrate hindgut // Nature 565 (7740), 480–484 (2019). doi 10.1038/s41586-018-0865-9 {Evarts2019} Holly Evarts. How Stem Cells Self-Organize in the Developing Embryo // Press-Relis Новое исследование использует визуализацию в реальном времени, чтобы понять критический шаг в раннем эмбриональном развитии - как гены и молекулы контролируют силы, управляющие появлением формы в развивающемся эмбрионе. 16/1/2019 {Free2019} Tristan Free. Cellular organization in embryonic development: it’s all in the concentration // BioTechniques 29/1/2019 Peter Belobrov 02 Feb 2019 12:02 PIT00448 KEGG & Cell Book – взаимное действие KEGG: Kyoto Encyclopedia of Genes and Genomes — веб-ресурс, предоставляющий доступ к ряду биологических баз данных и инструментам для анализа биологических и медицинских данных, созданный в 1995 году в рамках проекта «Геном человека». С момента создания интегрированная база данных KEGG значительно расширилась и на данный момент (2017) насчитывает шестнадцать баз данных, для удобства поиска разделенных на четыре категории: системная информация (данные о метаболических путях, модулях генов и т. д.), геномная информация (данные об отдельных генах, геномах различных существ, ортологичных группах), химическая информация (данные о химических реакциях, происходящих в живых организмах) и информация, связанная непосредственно со здоровьем человека (данные о человеческих болезнях, лекарствах и т. д.). Также KEGG предоставляет ряд инструментов для удобной работы с базами данных и анализа хранящейся в них информации. Идея Ильи Потапенко о красоте и важности Киотской энциклопедии генов и геномов для нашего проекта CellBook + Книга клетки + Cell Book была высказана почти 2 года назад на канале #cellbook 7/4/2017. Но идея не захватила массы сразу, к сожалению! Теперь просматривая Энциклопедию в целом обращаю Ваше внимание, что хорошим путеводителем по ней является статья, которая входит в число добротных статей русскоязычного раздела Википедии KEGG. Кстати, это, действительно, добротная статья, которая намного более английского аналога в Wiki, и которая за разумное время позволяет Вам понять суть энциклопедии. Давнее моё желание перестать пользоваться в биологии словом "взаимодействие = interaction", заменяя его везде, где это точно определено, на "биологическое действие = biological action", очень выпукло иллюстрируют прогулки по Киотской энциклопедии генов и геномов. Удачи Вам всем в научных поисках. Peter Belobrov 31 Jan 2019 07:20 PIT00414 Секреты тотипотентных клеток Как тотипотентная клетка умудряется сохранить потенциал, чтобы стать любой из более чем 200 типов клеток в организме человека, является давней загадкой в биологии. Словами Тимофеева-Ресовского это ключевой момент основной биологической проблемы "как проблемы передачи наследственной информации от поколения к поколению и реализации этой информации в каждом поколении. В сущности, такая формулировка покрывает всю основную проблематику биологии" (источник и точную цитату см в PIT00356). Существенное продвижение в разгадке этого секрет сделано в недавно опубликованной работе [Flyamer2017]. Для понимания отношения этого поста к родословной клеток полезно посмотреть более ранние посты PIT00069, PIT00070, PIT00220 и PIT00259. В статье [Flyamer2017] среди авторов исследователи из команды Сергея Владимировича Разина. Без хотя бы одноразового чтения книги С. В. Разин, А. А. Быстрицкий. Впервые в мире продемонстрировано, что фрагмент ДНК, картированный в качестве петли с использованием биохимического подхода, действительно образует петлю, которая может быть визуалирована на цитологических препаратах. На примере домена альфа-глобиновых генов кур изучены основные принципы работы геномных доменов открытого типа, которые не имеют четких границ и могут содержать перекрывающиеся ткане-специфичные гены и гены домашнего хозяйства. Обоснована гипотеза, постулирующая, что эволюция генома может происходить посредством делеций, дупликаций и перемещений топологических петель ДНК. Для более популярного изложения сути дела полезен пресс-релиз Института молекулярной биотехнологии Академии наук Австрии "Уникальная архитектура генома после оплодотворения в одноклеточный эмбрионах", из которого взята картинка с подписью ниже.  Слияние яйца и спермы, двух высокодифференцированных типов клеток, приводит к образованию одноклеточного эмбриона или зиготы. В первые часы после оплодотворения два отдельных генома подвергаются событиям перепрограммирования, которые предположительно функционируют, чтобы стереть память о дифференцированном типе клеток и установить состояние тотипотентности. Механизмы, лежащие в основе тотипотентности, остаются малоизученными, хотя они обязательны для создания нового организма из оплодотворенной яйцеклетки. {Flyamer2017} Ilya M. Flyamer, Johanna Gassler, Maxim Imakaev, Hugo B. Brandão, Sergey V. Ulianov, Nezar Abdennur, Sergey V. Razin, Leonid A. Mirny, and Kikuë Tachibana-Konwalski. Single-nucleus Hi-C reveals unique chromatin reorganization at oocyte-to-zygote transition // Nature 544 (7648), 110–114 (2017). doi 10.1038/nature21711 Аннотация. Хроматин перепрограммируется после оплодотворения для получения тотипотентной зиготы с потенциалом для создания нового организма [1]. Материнский геном, унаследованный от ооцита, и отцовский геном, обеспечиваемый спермой, сосуществуют как отдельные гаплоидные ядра в зиготе. То, как эти два эпигенетически обособленных генома пространственно организованы, изучено плохо. Существующие методы фиксации конформации хромосом [2, 3, 4, 5] не применимы к ооцитам и зиготам из-за недостатка материала. Чтобы исследовать трехмерную организацию хроматина в редких типах клеток, мы разработали протокол одноядерного захвата конформации хромосом высокого разрешения (snHi-C), который обеспечивает более чем в 10 раз больше контактов на клетку, чем предыдущий метод [2]. Здесь мы показываем, что архитектура хроматина уникально реорганизуется во время перехода ооцит-зигота у мышей и отличается в отцовских и материнских ядрах в одноклеточных зиготах. Особенности геномной организации, включая компартменты, топологически ассоциирующие домены (TAD) и петли, присутствуют в отдельных ооцитах при усреднении по геному, но наличие каждого признака в локусе варьируется между клетками. На уровне менее миллиона оснований (sub-megabase) мы наблюдали стохастические кластеры контактов, которые могут встречаться через границы TAD, но в среднем в TAD (см PIT00259). Примечательно, что мы обнаружили, что TAD и петли, но не компартменты, которые присутствуют в материнском хроматине зиготы, предполагая, что они генерируются различными механизмами. Наши результаты показывают, что глобальная организация хроматина ядер зиготы принципиально отличается от структуры других интерфазных клеток. Понимание этого основного состояния хроматина зиготы может потенциально обеспечить понимание перепрограммирования клеток в состояние тотипотентности. 1. Zhou, L. Q. & Dean, J. Reprogramming the genome to totipotency in mouse embryos. Trends Cell Biol. 25, 82–91 (2015) 2. Nagano, T. et al. Single-cell Hi-C reveals cell-to-cell variability in chromosome structure. Nature 502, 59–64 (2013) 3. Ma, W. et al. Fine-scale chromatin interaction maps reveal the cis-regulatory landscape of human lincRNA genes. Nat. Methods 12, 71–78 (2015) 4. Hsieh, T.-H. S. et al. Mapping nucleosome resolution chromosome folding in yeast by micro-C. Cell 162, 108–119 (2015) 5. Barutcu, A. R. et al. C-ing the genome: a compendium of chromosome conformation capture methods to study higher-order chromatin organization. J. Cell. Physiol. 231, 31–35 (2016) Peter Belobrov 21 Apr 2017 02:20 Продолжая пост PIT00361, PIT00362 и PIT00363 рекомендую почитать статью [Trcek2017], в которой вы найдёте разгадки нескольких тайн «зародышевых или P гранул». Вот рисунок из статьи [Trcek2017], давшей название этого поста. 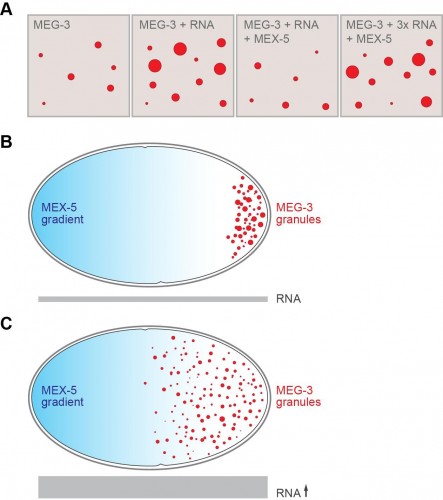 Рис.1 [Trcek2017] {Trcek2017} Tatjana Trcek, and Ruth Lehmann. All about the RNA after all // eLife 6, e24106 (2017). doi 10.7554/eLife.24106 Peter Belobrov 25 Feb 2017 02:00 "Посмотрим правде в глаза", - пишут авторы статьи [Hallgrimsson2014], обсуждая проблемы создания моделей отношения генотип-фенотип в сложных признаках. Рассмотрим новые идеи проявления экспрессия генов в функциях организма опираясь на рисунок из этой статьи. 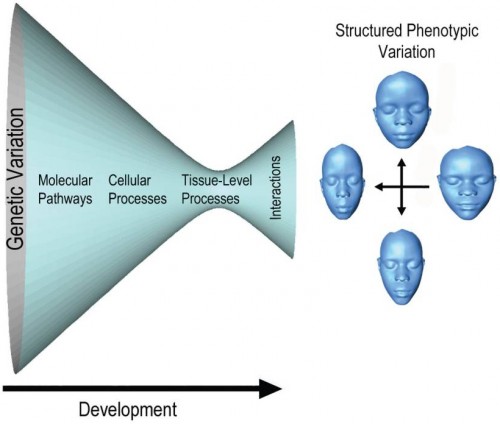 Обобщенный взгляд на факторы, влияющие на развитие лицевой структуры, отражает идею о том, что большое количество генетических вариаций направлено на меньшее количество путей и процессов [Hallgrimsson2014]. Противоположный взгляд на архитектуру развития, лежащей в основе отображения генотипа в фенотип для сложных признаков, рассмотрен со ссылкой на Вагнера [Wagner1996], показывает отображение генов «многие к многим» по фенотипическим признакам. Наша сказочка bioALgebr1 - это первая попытка разработать символьный биологический анализ, если угодно, в каком-то смысле, "биологическую алгебру" деления и развития клеток для проекта CellBook. Символьный анализ есть на Maple и MapleSim, однако это программное обеспечение инженерных расчетов с уже разработанной "инженерной алгеброй". Сразу после семинара обсуждение с Антоном "биологического умножения" дало некоторые варианты записи для "биологической алгебры". Чуть подробнее. В субботу 11/2/17 на семинаре как бы походя, была высказана простая мысль. Если у нас два типа связей, которые могут образоваться и разорваться, то у нас есть только физическая модель. Взяв полимер в растворители и даже несколько разных полимеров получаем химическую модель. И лишь когда мы возьмём распределение множественных связей со своим распределением по типам, энергиям и характерным расстояниям, то всё это уже становится биологической моделью, но только при том условии, что существует уровни ниже и уровни выше, на которых имеют место не менее простые, и даже более сложные типы взаимодействий. Конечно, без образа паттерна и процессов его передачи невозможно построить формальную биологическую модель. Но такие модели надо пытаться строить, чтобы создать некоторые канонические модели, преобразования в которых будут определять "биологическую алгебру". Антон спросил о биологическом умножении. Ответ опять-таки родился сам собой. Умножение (множенные клетка) вместе с делением и приростом (или без - чистое дробление) массы и увеличением разнообразия типов экспрессии. Биологи называют это развитием. То, что предложено выше - это одна из попыток построить биологическую модель развития. Необходимо подчеркнуть важность беспокойных состояний и процессов созидания нового. Приведу здесь вчерашнее обсуждение сказочки bioALgebr1 с Иваном для проекта CellBook. Берем два типа объектов и несколько типов связей, чтобы изучать некоторые канонические преобразования их массы. [9:51] [9:53] [9:54] [9:55] [9:59] [10:13 PM] [10:15 PM] Это всё о позволило утром родиться следующей мысли. MoLLibs, MoLBooks, ChemLibs, ChemBooks, CellBooks, CellLibs - это те образы, точнее, прообразы, функции которых создают паттерны вместе с уровнями организации. Важная находка для проекта CellBook. Множества этих клякс могут образовать читалки, писалки, преобразовалки … да и распределения их – важный объект взаимных действий. Кстати, чтение и запись – это функции вполне определенных множеств или их объединений. Важно подчеркнуть. Это не математические множества с одинаковыми элементами, но биологические множества, все элементы которого разные, хотя может существовать некоторая эквивалентность по группам элементов. {Hallgrimsson2014} Benedikt Hallgrimsson, Washington Mio, Ralph S. Marcucio, and Richard Spritz. Let's face it—complex traits are just not that simple // PLoS genetics 10 (11), e1004724 (2014). doi 10.1371/journal.pgen.1004724 {Wagner1996} Günter P. Wagner. Homologues, natural kinds and the evolution of modularity // American Zoologist 36 (1), 36-43 (1996). doi 10.1093/icb/36.1.36 Peter Belobrov 14 Feb 2017 09:00 PIT00385 Гибрид человека и свиньи Такие «страсти-мордасти» наводят «тень на плетень». Невольно вспоминаешь пословицу «если человеку долго говорить, что он - свинья, то человек захрюкает». Реферат статьи [Wu2017] с обсуждением проблемы создания гибрида человека и свиньи Е. Ефимова. Создан гибрид человека и свиньи. Каковы преимущества и риски спорной методики? // vesti.ru 27/1/2017 позволил обратиться к оригинальным статьям [Mascetti2016, Wu2016, Wu2017]. Экспериментальные химеры широко признаны в качестве самых строгих тестов для проверки плюрипотентности стволовых клеток. Предимплантационная химеры обеспечивают клетки-доноры с доступом развития для всего плода и внеэмбриональной мезодермой (желтка, аллантоиса и амниотической мезодермы), тем самым обеспечивая широкую оценку потенциала развития донорских клеток. Тетраплоидные предимплантационная химеры, в частности, считаются наиболее полным испытанием плюрипотентности, потому что из стволовых клеток полученных полностью потомство мыши, что являются конечной точкой оценки [Mascetti2016]. Химеры являются монстрами воображения и давно установившимся исследовательским инструментом. Последние достижения, особенно те, которые касаются идентификации и генерации различных видов стволовых клеток, расширили репертуар и полезность межвидовых химер млекопитающих и привели к новым путям пониманию фундаментальной биологии и потенциальных клинических приложений [Wu2016]. Понятно, что на самом деле никакого гибрида человека и свиньи нет и не будет. Создана химера из плюрипотентных клеток человека и свиньи. Графическая аннотация ниже поясняет суть работы [Wu2017]. 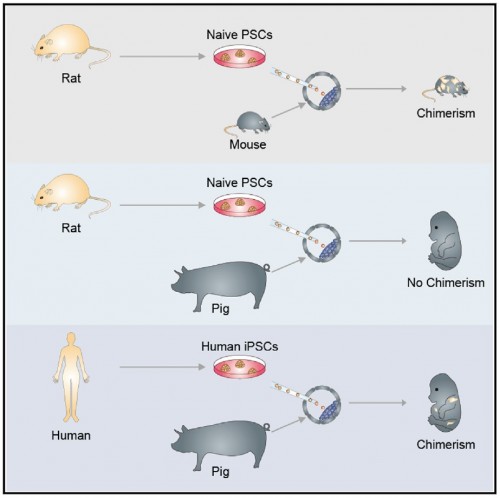 Человеческие плюрипотентные стволовые клетки хорошо приживаются в предимплантационные бластоцисты крупного рогатого скота и свиней, но показывают ограниченный химерный вклад в постимплантационные свиные эмбрионы. Это направление очень активно развивается для понимания биологических взаимодействий между клетками и тканями. Вот 3 самые последние работы для понимания состояния проблемы {Wu2017} Jun Wu et al. Interspecies Chimerism with Mammalian Pluripotent Stem Cells // Cell 168 (3), 473–486.e15 (2017). doi 10.1016/j.cell.2016.12.036 {Mascetti2016} Victoria L. Mascetti, Roger A. Pedersen. Contributions of mammalian chimeras to pluripotent stem cell research // Cell Stem Cell 19 (2), 163-175 (2016). doi 10.1016/j.stem.2016.07.018 {Wu2016} Jun Wu et al. Stem cells and interspecies chimaeras // Nature 540 (7631), 51-59 (2016). doi 10.1038/nature20573 Peter Belobrov 29 Jan 2017 12:00 PIT00382 CRISPR и танцы молекул Вчера вместо Крещенского купания получил по почте новую удивительно интересную и заставляющую думать книгу Дениса Нобла "Танцы под дудочку жизни. Биологическая относительность". Denis Noble. "Dance to the tune of life. Biological relativity". Cambridge University Press. 2017 В этой книге формулируется теория биологической относительности, подчеркивающая, что живые организмы работают на разных уровнях сложности и, следовательно, должны быть проанализированы на многих масштабах с релятивистской точки зрения. Нобл объясняет, что все биологические процессы протекают в молекулярных, клеточных и организменных сетях. Интерактивный характер этих фундаментальных процессов лежит в основе биологической теории относительности, как таковой, и проблемы упрощенного молекулярного редукционизма. Нобл показывает, что такой интегративный подход возникает как необходимое следствие строгого применения математики к биологии. Опираясь на свою новаторскую работу в математической физике биологии, он показывает, что появление этого является глубоко гуманной картиной роли организма в сдерживании его химии, в том числе его генов, чтобы служить всему организму в целом, особенно при взаимодействии с его социальной средой. Эта гуманистический, целостный подход ставит под сомнение общепринятые представления о генах, которые находятся в центре, и которых придерживаются многие в современной биологии и в культуре. Денис Нобл является заслуженным профессором физиологии сердечно-сосудистой системы и директором центра вычислительной физиологии Оксфордского университета. Он является президентом Международного союза физиологических наук и членом Королевского общества, Великобритания. Краткое содержание книги "Танцы под дудочку жизни" 1 • Вселенная и принцип относительности 2 • Биологические масштабы и уровни 3 • Биологические сети 4 • Природа и происхождение клеток 5 • Слепой случай и естественный отбор 6 • Биологическая теория относительности 7 • Танцы нуклеотидов: природная генная инженерия 8 • Эпигенетика и релятивистская теория эволюции 9 • Относительность эпистемологии: Значение всего этого Смысл названия главы 9 напоминает слова Владимира Высоцкого "Ведь даже Эйнштейн - физический гений - весьма относительно всё понимал". Буду читать и пытаться найти переводчика, так как первая книга Дениса Нобла "Музыка жизни. Биология за пределами генома", вышедшая 10 лет назад, так и не была переведена на русский язык, хотя были сделаны переводы на многие языки мира. Подробнее см. на сайте musicoflife. Denis Noble. "The Music of Life. Biology beyond the Genome". 2006. 176 p. На нашем сайте более десятка постов основаны на "Музыке жизни". Проблемы природной генной инженерии связаны с танцами нуклеотидов (Гл.7) в книге "Танцы под дудочку жизни". Важным примером природной генной инженерии является CRISPR, см., например, {Mikuni2016} Takayasu Mikuni, Jun Nishiyama, Ye Sun, Naomi Kamasawa, and Ryohei Yasuda. High-throughput, high-resolution mapping of protein localization in mammalian brain by in vivo genome editing // Cell 165 (7), 1803-1817 (2016). doi 10.1016/j.cell.2016.04.044 Так как книга в России ещё мало доступна, рекомендую прочитать статью [Mikuni2016] вместе со статьёй [Noble2013], чтобы "навести на резкость" понимание современного взгляда на проблему молекулярных танцев под дудочку жизни в технологии CRISPR. {Noble2013} Denis Noble. A biological relativity view of the relationships between genomes and phenotypes // Progress in biophysics and molecular biology 111 (2), 59-65 (2013). doi 10.1016/j.pbiomolbio.2012.09.004 Peter Belobrov 20 Jan 2017 04:44 Вот мы и дождались новость, которую считали очевидной с точки зрения CellBook и родословной клетки. В поле эпигенетики обнаружена важная новая функцию RNAi. Вот основная статья {Roche2016} B. Roche, B. Arcangioli, and R. A. Martienssen. RNA interference is essential for cellular quiescence // Science 354 (6313), aah5651 (2016). DOI 10.1126/science.aah5651 Вот рисунок из структурированной аннотации этой статьи 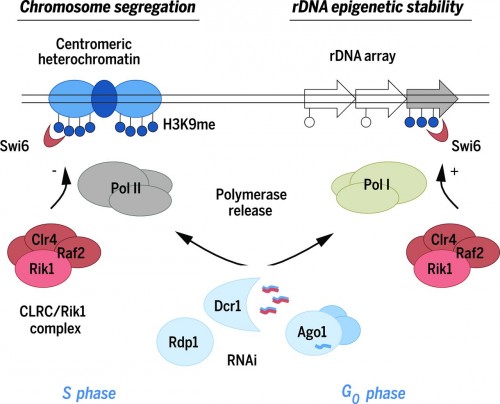 [Roche2016] Двойная роль РНК-интерференции в фазах покоя S и G0 клеточного цикла на основе освобождения РНК-полимеразы. Выделение РНК Pol II позволяет формировать гетерохроматин на центромерах, и выделение РНК Pol I из рДНК позволяет избежать переизбытка гетерохроматина во время фаз покоя. В нижней части рисунка, красные и синие изогнутые линии представляют собой малые РНК. Выводы основываться на классическом исследовании группы Мартинсена, где было показано, что удаление генов, кодирующих компоненты путей RNAi в делящихся дрожжах приводит к потере молчания генов [Volpe2002]. {Volpe2002} Thomas A. Volpe, Catherine Kidner, Ira M. Hall, Grace Teng, Shiv IS Grewal, and Robert A. Martienssen. Regulation of heterochromatic silencing and histone H3 lysine-9 methylation by RNAi // Science 297 (5588), 1833-1837 (2002). DOI 10.1126/science.1074973 Cited by 1869 Работа, проведенная в лаборатории Мартинсена, проливает свет на клеточное состояние, которое не учитывается в большинстве молекулярных исследований. Это сказано в развернутом реферате "Интерференция РНК держит клетки в покое" статей [Roche2016] и [Volpe2002]. Графическая аннотация из этого реферата приведена ниже для понимания фаз S и G0 клеточного цикла 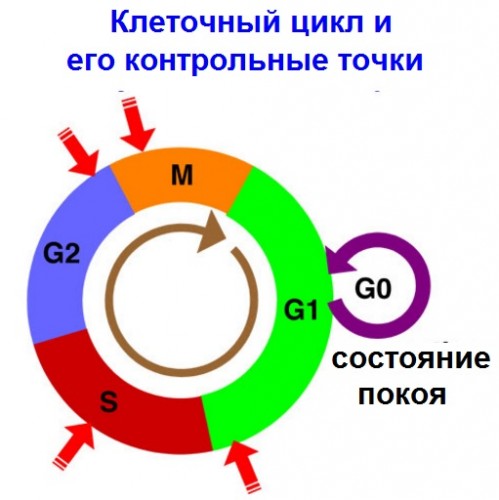 Научным руководителем исследований, результаты которых - тема этого поста, является Роберт А. Мартинсен (Robert A. Martienssen) - профессор в области генетики растений из Колд Спринг Харбора (Cold Spring Harbor). Он является экспертом по метилирования заменяемых элементов (транспозонов) в растительных геномах. Он также исследовал связь между метилированием ДНК, метилированием гистонов и РНК-интерференцией в растениях и дрожжах. Область его интересов - перепрограммирование гетерохроматина, малые интерферирующие РНК и судьба зародышевых клеток. Важность этих результатов подчеркивается в недавнем обзоре "Некодирующие РНК как регулятор в эпигенетике", для которого пока доступна лишь аннотация Jian-Wei Wei, Kai Huang, Chao Yang, and Chun-Sheng Kang. Non-coding RNAs as regulators in epigenetics (Review) // Oncology Reports 37 (1), 3-9 (2017). DOI 10.3892/or.2016.5236 Peter Belobrov 08 Dec 2016 08:00 PIT00368 Микробиомы разные Микробиомы не просто разные! Действие антибиотиков на микрофлору кишечника и обмен веществ зависит от принимающего антибиотики. Действие антибиотиков очень индивидуально и сильно зависят от генетики человека. Это показано в недавней работе {Fujisaka2016} Shiho Fujisaka, Siegfried Ussar, Clary Clish, Suzanne Devkota, Jonathan M. Dreyfuss, Masaji Sakaguchi, Marion Soto et al. Antibiotic effects on gut microbiota and metabolism are host dependent // The Journal of Clinical Investigation 126 (12), 4430-4443 (2016). doi 10.1172/JCI86674 В расширенном реферате один микробиомом не подходит для всех популярно рассмотрена результаты статьи [Fujisaka2016]. Заменю пересказ простым напоминанием, того, что когда едите, не забывайте о родословной, т.к. точной характеристики связи результатов действия антибиотиков, ожирения и диабета с генетическим паспортом пока не удалось установить. Хотя зависимость от генетики в работе [Fujisaka2016] доказана. Тот факт, что в иммунной системе все микробиомы разные хорошо известен. А вот что хорошо для одного, может быть плохим для другого в зависимости от родословной, с этим нельзя не считаться. Пока не ясна генетика нельзя действовать на микробиом одними и теми же лекарствами. Действительно, индивидуальная медицина нужна. А вот насколько микробиомы разные? Чтобы не расслабляться, задайтесь вопросом, как микробиомы влияют друг на друга. Для того, чтобы исследовать, как микробиом адаптируется к новой среде, обратим внимание на приматов. В работе [Clayton2016] изучена микрофлора из двух видов обезьян, живущих в зоопарке, в храме, или в дикой природе. Данные показали, что у обезьян, живущих в дикой природе широкое разнообразие микробиомов, в то время как животные, живущие в святилищах показали значительно меньшее разнообразие. Интересно, что обезьяны, живущие в зоопарках, показали еще меньше разнообразия, а их микробиомы на самом деле напоминают найденные у современных людей. Последующее исследование 33 обезьян, представляющих 8 различных видов, взятые из зоопарков показали аналогичную микробную комплементарность. {Clayton2016} Jonathan B. Clayton, Pajau Vangay, Hu Huang, Tonya Ward, Benjamin M. Hillmann, Gabriel A. Al-Ghalith, Dominic A. Travis et al. Captivity humanizes the primate microbiome // Proceedings of the National Academy of Sciences 113 (37), 10376–10381 (2016). doi 10.1073/pnas.1521835113 Peter Belobrov 06 Dec 2016 02:38 PIT00363 Капельные органеллы - глоссарий Капельные органеллы, рассмотренные в статье [Li2012] и обзоре [Courchaine2016] - это, пожалуй, и есть капельные сборки, о которых мы не раз говорили. Почти 3 месяца спустя возвратимся сюда для подробного чтения, т.к. появилась статья [Smith2016], где активно используется [Courchaine2016] для обоснования и объяснение экспериментов. Стало понятно, что работа [Courchaine2016] является классикой, где клеточные сборки рассматриваются на языке разделения фаз жидкость-жидкость или LLPS. [Smith2016] начинает текст статьи фразой: «РНК гранулы образуются концентрирующей сборкой РНК и РНК-связывающих белков, которые формируют гранулы (капли) без ограничивающей мембраны в цитоплазме или нуклеоплазме клеток [Courchaine2016]. РНК гранулы вездесущие клеточные структуры. Несколько классов цитоплазматических гранул РНК были описаны, в том числе стрессовые гранулы, P тельца, гранулы в нейронах и зародышевые гранулы». Полезен глоссарий из [Courchaine2016]. Разделение фаз жидкость-жидкость (LLPS) Домены низкой сложности (LCD) Внутренне разупорядоченные области (IDRs) Капелька Гидрогель Агрегаты Амилоид Прион-подобные домены Не очень понятно, почему низкой сложностью называют нерегулярные случайные пространственные структуры полипептидов? С точки зрения капельного направления разделения фаз жидкость-жидкость в больших каплях, пожалуй, стоит принять во внимание. Например, традиционные представления о равновесии микро- и макроэмульсий складывались на основе взаимодействия масла с водой и на формировании сверхнизкого поверхностного натяжения за счёт ПАВ и соПАВ (спирты). Слабо упорядоченные белки вместе с РНК образуют гранулы с другим типом фазового расслоения жидкость-жидкость и позволяют в клетке формировать гранулы микронных размеров. {Li2012} Pilong Li, Sudeep Banjade, Hui-Chun Cheng, Soyeon Kim, Baoyu Chen, Liang Guo, Marc Llaguno et al. Phase transitions in the assembly of multivalent signalling proteins // Nature 483 (7389), 336-340 (2012). doi 10.1038/nature10879 Cited by 288 20/1/2026 Cited by 2790 {Courchaine2016} Edward M. Courchaine, Alice Lu, and Karla M. Neugebauer. Droplet organelles? // The EMBO journal 35 (15), 1603–1612 (2016). DOI 10.15252/embj.201593517 Cited by 14 20/1/2026 Cited by 333 {Smith2016} Jarrett Smith, Deepika Calidas, Helen Schmidt, Tu Lu, Dominique Rasoloson, and Geraldine Seydoux. Spatial patterning of P granules by RNA-induced phase separation of the intrinsically-disordered protein MEG-3 // eLife 5, e21337 (2016). doi 10.7554/eLife.21337 20/1/2026 Cited by 263 Peter Belobrov 01 Dec 2016 18:34 PIT00362 Вернёмся к сборкам в клетке Чтобы понять нашу точку зрения на молекулярные Книги Клетки (CellBook), вернёмся к посту PIT00124, где был сделан подробный реферат статьи [Toretsky2014]. Глубина изменения представлений о связи структуры и функции белков видна в Белковой Троице, обобщенную авторами [Dunker2001] в работе [Dunker2001b]. 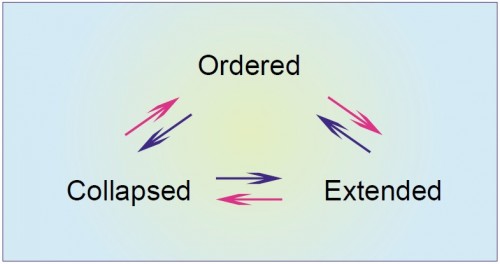 Рис. 1 [Dunker2001b]. Белковая Троица. Нативные белки могут существовать в одном из трех состояний - упорядоченном, разрушенном неупорядоченном, или расширенном неупорядоченном. Словосочетание "биологическая сборка" употребляют достаточно часто. Вот новые ссылки [Uversky2017] и др. на работу [Toretsky2014], которая очень интересны для обсуждения сборок в клетках. Начало было положено статьёй [Uversky2002] «Нативно развернутые белки: точка, в которой биология ждет физики». В работе введено состояние предрасплавленной глобулы и показано, что полипептиды без упорядоченной структуры при физиологических условиях являются общим правилом, а не исключением. Ниже приведена суммирующий рисунок из этой работы. 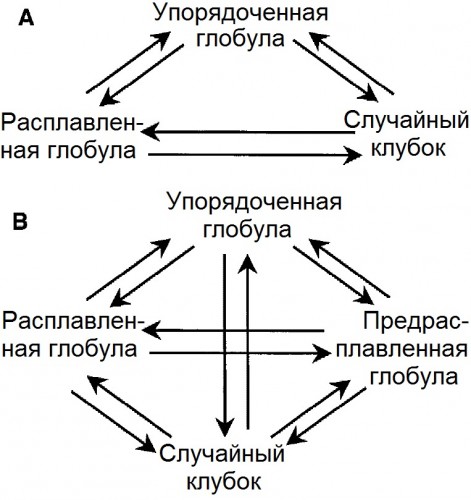 Рис. 6 [Uversky2002]. Расширение Белковой Троицы [Dunker2001] (А) до Белкового Квартета модели функционирования белка (B). В соответствии с этой моделью, функция возникает из четырех специфических конформаций полипептидной цепи (упорядоченные формы, расплавленные глобулы, предрасплавленные глобулы и случайные клубки) и переходов между любыми из этих состояний. Развитие точки зрения В.Н. Уверского (школа О.Б. Птицина, Институт белка РАН) за 15 лет на белковые сборки в клетках можно понять на приведенных ниже рисунках, где введены чисто биологические понятия: посттрансляционной модификации (PTMs), белковых органелл без мембран (PMLOs), которые образуются в результате расслоения жидкой смеси (также известной как фазовый переход жидкость-жидкость, LLPT) или коацервации, которая является одной из форм конденсации в водных растворах белков. (PMLOs) 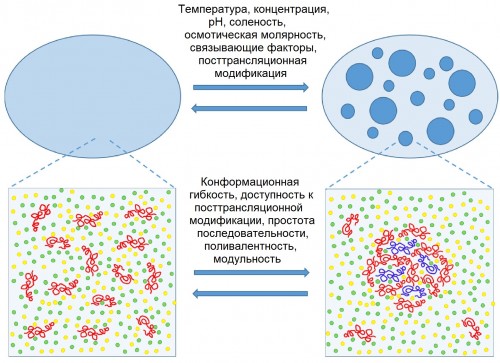 Рис.1 [Uversky2017]. Термодинамические факторы (вверху) и признаки беспорядка контролирующие фазовые переходы жидкость - жидкости в растворах белков. 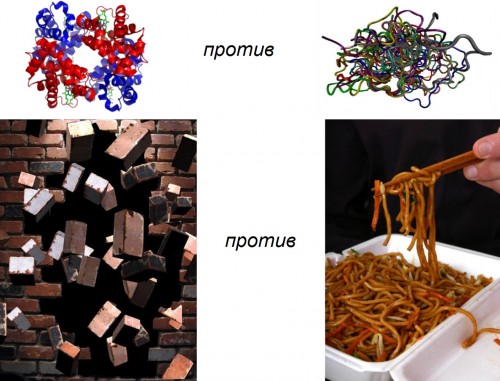 Рис.2 [Uversky2017]. Собственный беспорядок и устойчивость разделенных фаз капель. Упорядоченные и собственно неупорядоченные белки, представленные на этих рисунках лишь модели, которые не связаны с белковыми органеллами без мембран (PMLOs). А сейчас хочу обратить ваше внимание на то, что молекулярные сборки в клетках включают не только белки, но ещё РНК, ДНК, липиды и множество других молекул, выполняющих биологические функции. {Dunker2001} AK Dunker et al. Intrinsically disordered protein // J Mol Graph Model. 19 (1), 26-59 (2001). PMID 11381529 Cited by 1848 {Dunker2001b} A. Keith Dunker, and Zoran Obradovic. The protein trinity—linking function and disorder // Nature biotechnology 19 (9), 805-806 (2001). doi 10.1038/nbt0901-805 Cited by 502 {Uversky2002} Vladimir N. Uversky. Natively unfolded proteins: a point where biology waits for physics // Protein science 11 (4), 739-756 (2002). DOI 10.1110/ps.4210102 Cited by 1433 Аннотация. Сделан анализ экспериментального материала, накопленного в литературе по конформационному поведению внутренне неструктурированных (изначально развернутых) белков. Результаты этого анализа показали, что эти белки не обладают равномерными структурными свойствами, как можно ожидать для членов единого термодинамического семейства. Скорее всего, эти белки могут быть разделены на две структурно различные группы: истинные клубки и предрасплавленные глобулы. Белки из первой группы имеют гидродинамические размеры, характерные для случайных клубков в плохом растворителе и не обладают какой-либо (или почти любой) определённой вторичной структурой. Белки из второй группы, по существу, более компактные, обнаруживают некоторое количество остаточных вторичных структур, хотя они менее плотные, чем исходные или расплавленной глобулы белков. Важной особенностью внутренне неструктурированных белков является то, что они претерпевают переход порядок-беспорядок во время или перед выполнением их биологической функции. В связи с этим обсуждается модель "Белкового Квартета", с функцией, вытекающих из четырех конкретных конформаций (упорядоченные формы, расплавленные глобулы, предрасплавленные глобулы и случайных клубков) и переходов между любыми двумя состояниями. {Toretsky2014} J A Toretsky & P E Wright. Assemblages - Functional units formed by cellular phase separation // JCB, 206 (5), 579-588 (2014). DOI 10.1083/jcb.201404124 Cited by 57 {Uversky2017} Vladimir N. Uversky. Intrinsically disordered proteins in overcrowded milieu: Membrane-less organelles, phase separation, and intrinsic disorder // Current Opinion in Structural Biology 44, 18-30 (2017). doi 10.1016/j.sbi.2016.10.015 {Uversky2017b} Vladimir N. Uversky. How to Predict Disorder in a Protein of Interest // Prediction of Protein Secondary Structure (2017), 137-158. doi 10.1007/978-1-4939-6406-2_11 Аннотация. Доступные в настоящее время вычислительные инструменты, которые много, обеспечивают исследователю множество вариантов для прогнозирования внутреннего беспорядка у белка, представляющего интерес, и для обнаружения по меньшей мере, некоторые из его беспорядков, связанных с основными функциями. В этой главе дается весьма субъективный обзор того, как не потеряться в «темном лесу» доступных инструментов для анализа внутреннего беспорядка. Но он никоим образом не дает уникального пути через этот лес, а просто представляет некоторые из инструментов, которые автор использует в своих повседневных исследованиях. Peter Belobrov 01 Dec 2016 14:14 Отмечая 4 года посту PIT00003 P гранулы Вариант 2, 2 года - PIT00124 и год - PIT00207, хочу обратить ваше внимание на принципиально новый взгляд на комплексы РНК с белком. Этому мы обязаны в значительно степени лаборатории Дженнифер Додны (Jennifer A. Doudna), на 1-й странице сайта которой бактериальный иммунитет, РНК интерференция эукариот и контроль трансляции. Это, пожалуй, три основных инструмента книги клетки (CellBook), которые открыты в лаборатории Дженнифер. B.T. Staahl, M. Benekareddy, C. Coulon-Bainier, A. Ghosh, J.A. Doudna. Efficient protein-based genome editing by local delivery in the brain // Poster 317.09/T7 at 2016 Neuroscience Meeting, Session 317 - Neuroprotective Mechanisms: Signaling and Gene Expression of 14/11/2016. Abstract. RNA-guided genome editing triggered by the CRISPR-Cas9 DNA endonuclease has the potential to cure the underlying cause of a genetic disease. This therapeutic strategy will require tissue-specific delivery of the editing molecules in vivo, a goal that is particularly challenging in the brain. Here we show the Cas9 ribonucleoprotein (RNP) complexes can be used to edit post-mitotic neurons in the mouse brain. We also show that protein engineering of Cas9 increased the efficiency of in vivo neuronal cell editing ten-fold. Cas9 RNPs introduced locally in the brain trigger genome editing in cells including hippocampal, striatal and cortical neurons by a mechanism that involves spreading beyond the immediate site of injection. We also show that Cas9 RNP complexes are stable over 6 hours in blood plasma. These advances provide a robust technology for application of genome-editing in the brain to treat the underlying cause of neurological genetic diseases. Аннотация. РНК-управляемое редактирование генома инициируется CRISPR-Cas9 эндонуклеазами ДНК, которые имеет потенциал для лечения основной причины генетического заболевания. Эта терапевтическая стратегия требует доставки тканеспецифических молекул для редактирования in vivo. Эта цель является особенно сложной в головном мозге. Показано, что Cas9 рибонуклеопротеиновый (Cas9 RNP) комплексы могут быть использованы для редактирования пост-митотических нейронов в головном мозге мыши. Авторы также показали, что Cas9 белковая инженерия повысила эффективность редактирования нейронов in vivo в десять раз. Cas9 RNPs вводились локально для редактирования геномов переключающих клеток мозга, включая гиппокамп, полосатое тело и кортикальные нейроны по механизму, который включает в себя распространение за пределы непосредственного места инъекции. Авторы также показали, что Cas9 RNP комплексы стабильны в течение 6 ч в плазме крови. Эти достижения обеспечивают надежную технологию для применения редактирования генома в мозге при лечении основной причины неврологических генетических заболеваний. И хотя ни постера ни статьи нет ещё, вчера эти тезисы прокомментировала Kayt Sukel. A Safe New Approach for CRISPR-Cas9 Gene Therapy in Humans // BioTechniques 29/11/2016 Вышел хороший обзор Аннотация. TBA (редактирую) А пока почитаем про механизм работы CRISPR-Cas9, который на редкость хорошо и аккуратно описан в русскоязычной Википедии CRISPR. Интересный для понимания сути дела текст на сайте биомолекула.ру CRISPR-системы - иммунизация прокариот. Более подробное рассмотрение можно найти в самых первых больших обзорах Patrick D. Hsu, Eric S. Lander, and Feng Zhang. Development and applications of CRISPR-Cas9 for genome engineering // Cell 157 (6), 1262-1278 (2014). doi 10.1016/j.cell.2014.05.010 Cited by 1074 Jennifer A. Doudna, and Emmanuelle Charpentier. The new frontier of genome engineering with CRISPR-Cas9 // Science 346 (6213), 1258096 (2014).DOI 10.1126/science.1258096 Cited by 798 Peter Belobrov 30 Nov 2016 04:06 PIT00357 7 полов инфузории тетрахимена Новость Как инфузория Tetrahymena thermophila выбирает себе пол была 3 года назад. У инфузории Tetrahymena thermophila семь полов. Вступить в половой процесс она может с представителем любого пола, кроме своего. Клетки, получившиеся в результате полового процесса, не наследуют пол у своих «родителей» — они выбирают его случайным образом из нескольких вариантов. Коллектив ученых из США и Китая в деталях разобрался в этом аспекте личной жизни инфузорий. Известно, что у всех инфузорий два генома — один, рабочий, содержится в макроядре, а второй в неактивной форме хранится в микроядре и предназначен для обмена генетической информацией. Ученые выяснили, что в макроядре за пол отвечает пара расположенных по соседству генов. В микроядре тетрахимены есть все варианты таких пар, но в укороченной форме. После полового процесса одна из них достраивается путем «склеивания» частей разных генов, а все остальные уничтожаются. Удивительно, что выбор пары, по-видимому, происходит случайным образом, но при этом требует очень аккуратной сборки фрагментов ДНК. После недавнего прочтения PIT00356 решил посмотреть новые ссылки на оригинальные статьи Marcella D. Cervantes, Eileen P. Hamilton, Jie Xiong, Michael J. Lawson, Dongxia Yuan, Michalis Hadjithomas, Wei Miao, and Eduardo Orias. Selecting one of several mating types through gene segment joining and deletion in Tetrahymena thermophila // PLoS Biol 11 (3), e1001518 (2013). doi 10.1371/journal.pbio.1002284 уже Cited by 37 Пока у меня нет собеседника. Поэтому перечитываю Тимофеева-Ресовского, который говорит о 10 полах. Помню мы с Виолеттой разбирали инфузории, у которых может быть порядка 1000 хромосом ... Кстати, даже из 16 000 хромосом состоит эукариотический геном инфузории Oxytricha trifallax. Estienne C. Swart, John R. Bracht, Vincent Magrini, Patrick Minx, Xiao Chen, Yi Zhou, Jaspreet S. Khurana et al. The Oxytricha trifallax macronuclear genome: a complex eukaryotic genome with 16,000 tiny chromosomes // PLoS Biol 11 (1), e1001473 (2013). doi 10.1371/journal.pbio.1001473 Cited by 66 В этом соматическим геноме находится более 16 000 уникальных нанохромосом ("nanochromosomes"), которые обычно несут единичные гены и имеют средний размер всего 3,2 кб. Термин нанохромосома (nanochromosome) был введен в 2003 в работе Thomas G. Doak, Andre RO Cavalcanti, Nicholas A. Stover, Diane M. Dunn, Robert Weiss, Glenn Herrick, and Laura F. Landweber. Sequencing the Oxytricha trifallax macronuclear genome: a pilot project // Trends in Genetics 19 (11), 603-607 (2003). doi 10.1016/j.tig.2003.09.013 Cited by 39 Через 10 лет появляется эта работа {Bracht2013} John R. Bracht, Wenwen Fang, Aaron David Goldman, Egor Dolzhenko, Elizabeth M. Stein, and Laura F. Landweber. Genomes on the edge: programmed genome instability in ciliates // Cell 152 (3), 406-416 (2013). doi 10.1016/j.cell.2013.01.005 Cited by 38 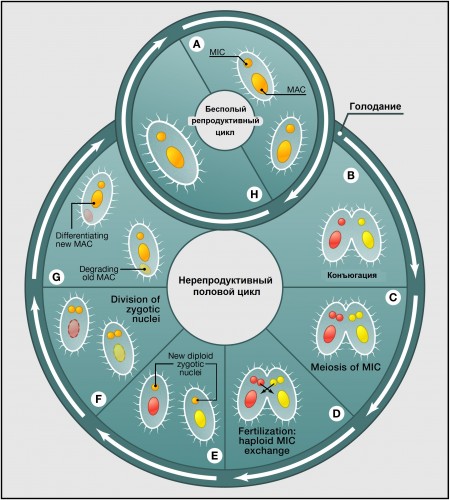 Рисунок 1. [Bracht2013] Упрощенные жизненные циклы ресничкообразных (А и Н) Репродуктивная, вегетативный рост происходит бесполым путем деления клеток, в том числе митозом зародышевой линии микроядер (MIC, обозначенной кружком) и амитозом соматического макронуклеуса (MAC, обозначается овалом). (B) Голодание индуцирует сопряжение между совместимыми типами спаривания, инициируя нерепродуктивный половой цикл. (C) Мейоз ВПК производит ядра гаплоидных гамет. (D и E) (D) Обмен гаплоидный микроядер и оплодотворение производит два новых диплоидных, зиготического ядра (Е). (F) Митоз зиготических ядер образуются два идентичных микроядра, и одно ядро начинает дифференцироваться в новый MAC. (G и H) (G) Деградация старого MAC происходит во время дифференцировки нового MAC. Зрелые клетки (H) снова вступают в вегетативный рост [Nowacki2009]. Для понимания приведу традиционную точку зрения. Конъюгация у инфузорий — обмен половыми ядрами (микронуклеусами) с последующим их попарным слиянием в синкарион (диплоидный продукт слияния миграционного и стационарного ядер у инфузории при конъюгации). Впоследствии синкарион делится с образованием новых половых и вегетативных ядер. {Nowacki2009} Mariusz Nowacki, Brian P. Higgins, Genevieve M. Maquilan, Estienne C. Swart, Thomas G. Doak, and Laura F. Landweber. A functional role for transposases in a large eukaryotic genome // Science 324 (5929), 935-938 (2009). DOI 10.1126/science.1170023 Cited by 93 В общем, дело пока проясняется. Понятно единственное, что наследование инфузориями родословной ещё преподнесёт много открытий! Например, уточнение понятия генома после того уже появился эпигеном ... Интересное переосмысливание термина геном, близкое к нашей книги клетки (CellBook), опубликовано недавно коллегами из Принстона, которые в этом году переехали в Колумбийский университет в Нью-Йорке. 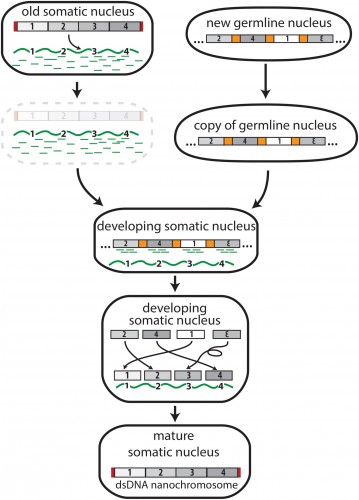 TBA figure caption in Russian Aaron David Goldman, and Laura F. Landweber. What Is a Genome? // PLoS Genet 12 (7), e1006181 (2016). doi 10.1371/journal.pgen.1006181 Аннотация. Геном часто описывается как хранилище информации организма. Является ли миллионы или миллиарды ДНК писем при их передаче через поколения основной средой для наследования организменных признаков. Некоторые развивающиеся области исследований показывают, что это определение является упрощением. Здесь мы рассмотрим способы, в которых более глубокое понимание геномного разнообразия и физиологии клетки является сложной задачей концепции физической неизменности, связанной с геномом, а также его роль в качестве единственного источника информации для организма. Цитата «С полным пониманием содержания генома, его разнообразия и способов выражения (экспрессии), теперь мы можем пересмотреть наше понимание генома и его роль в клетке. Например, при ближайшем рассмотрении определения NIH показывает, что две его половинки являются взаимоисключающими; то есть "полный набор ДНК" не может быть "всей информацией, необходимой для создания и поддержания взрослого(ых) организма." ... наше нынешнее понимание показывает, что это определение является неполным». Ещё цитата «Большая часть информации, необходимой для воспроизведения соматического генома происходит из РНК, а не из ДНК. Длинные, РНК-кэшируемые копии соматических хромосом от предыдущего поколения обеспечивают шаблоны для управления хромосомной перестройкой». Кроме этой статья коллеги из лаборатории Лоры Ландвебер доказали недавно утверждение «Архитектура зашифрованного генома показывает массивные геномные перегруппировки во время развития». Доказательство смотрите в работе Xiao Chen, John R. Bracht, Aaron David Goldman, Egor Dolzhenko, Derek M. Clay, Estienne C. Swart, David H. Perlman et al. The architecture of a scrambled genome reveals massive levels of genomic rearrangement during development // Cell 158 (5), 1187-1198 (2014). doi 10.1016/j.cell.2014.07.034 Cited by 33 Вот графическая аннотация этой статьи 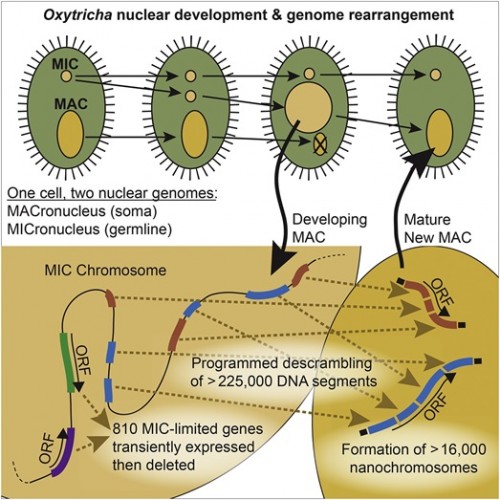 TBA figure caption in Russian Peter Belobrov 23 Nov 2016 15:46 PIT00356 Наследование родословной Тимофеев-Ресовский сказал, что «основную биологическую проблему, выражаясь современным языком, можно сформулировать так: это проблема передачи наследственной информации от поколения к поколению и реализации этой информации в каждом поколении. В сущности, такая формулировка покрывает всю основную проблематику биологии». Приведём его рассуждения про многополовость на стр. 26-27 в книге [1]: «Мы знаем, что, как правило, если отвлечься от исключений, жизнь на нашей Земле представлена формами, которым свойственно половое размножение с перекрестным оплодотворением и двуполовостью. Это вещь тривиальная, но я обращаю ваше просвещенное внимание на то, что и это удивительная и совершенно не сама по себе понятная вещь, потому что и на нашей планете в виде исключений мы имеем всякое: во-первых, бесполое размножение, во-вторых, вегетативное размножение, и, в-третьих, многополовость. Двуполовость, наличие двух полов — мужского и женского, есть некое эволюционное достижение на нашей планете. Мы еще до конца не понимаем, в строгой и ясной форме не показали, почему на нашей планете эволюция живых форм привела именно к дву-, а не к трех- или пятиполовости. Потому что мы знаем о существовании многополовости, в частности у ряда одноклеточных водорослей. Давно и в точной форме, в основном школами Книпа и Хартмана обнаружены виды, у которых имеется более десятка полов. 1. Peter Belobrov 22 Nov 2016 12:12 PIT00353 Молекулярные буквы и алфавиты Когда понимание того, что жизнь возможна только в органическом мире, и вся организация симбиозов направлена на сохранение органического материала, то мысль о том что каждая органическая молекула есть буква пришла само собой! Далее логично продолжать молекулярные слова, предложения, фразы ... короче до книги Клетки (CellBook) рукой подать. Об этом недавно написал Гудселл в своём видении молекулярных основ жизни David S. Goodsell. Atomic evidence: seeing the molecular basis of life. Springer, 2016. 180 p. Витзани также обсуждает эту проблему в статье Guenther Witzany. Crucial steps to life: From chemical reactions to code using agents // BioSystems 140, 49-57 (2016). А в препринте от 12/10/2016 Guenther Witzany. Two Genetic Codes: Repetitive Syntax for Active non-Coding RNAs; non-Repetitive Syntax for the DNA Archives // Preprint (2016). речь идёт о новом молекулярном синтаксисе для некодирующих РНК. Интересный поворот биологической логики! Peter Belobrov 20 Nov 2016 05:22 Продолжая рассмотрение родословной в рамках CellBook PIT00351, обратим внимание на наследование эпигенетики стволовых клеток, особенно не генетическое наследование и трансгенерационную эпигенетику. Понимание этой логики возможно лишь в допущении существования эпигенома на равных правах с геномом. В модели CellBook допускается более изящные варианты наследования родословной, независимые от генотипа, и дополняющие генетическое наследование. {Szyf2015} M. Szyf. Nongenetic inheritance and transgenerational epigenetics // Trends Mol. Med., 21, 134–144 (2015). doi 10.1016/j.molmed.2014.12.004 Аннотация. Идея, что наследственные генотипы определяют фенотипы имеет первостепенное значение в современной биологии. Остается открытым вопрос, могут ли стабильные фенотипы также унаследованы от родителей независимо от генетической последовательности. Последние данные свидетельствуют о том, что родительские переживания могут быть переданы поведенчески, через внутриутробное воздействие развивающегося плода в материнской среде, или через любой мужской или женской зародыш. Задача состоит в том, чтобы очертить вероятный механизм. За последнее десятилетие было предложено, что эпигенетические механизмы участвуют в передаче во многих поколениях фенотипов и в трансгенерационном наследования. Перспектива того, что переживания в родословной написаны в нашем эпигенома имеет огромные последствия для нашего понимания человеческого поведения, здоровья и болезни. Краткое изложение анализа статьи [Szyf2015] рассмотрено в статье {Sharma2015b} Abhay Sharma. Transgenerational epigenetic inheritance requires a much deeper analysis // Trends in molecular medicine 21 (5), 269-270 (2015). DOI 10.1016/j.molmed.2015.02.010 из которой мы приведём один рисунок. 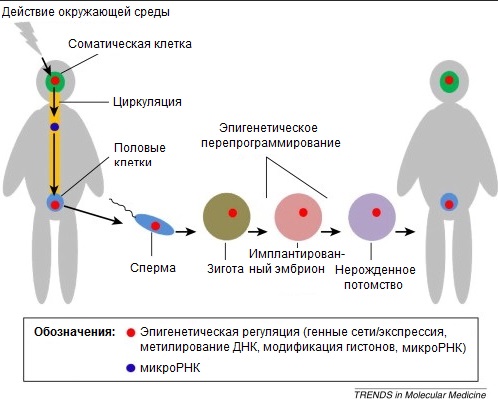 Рис. 1. Концептуальный, в рамках системной биологии трансгенерационная эпигенетическая наследственность. Экологический фактор индуцирует эпигенетические изменения в соматических клетках → циркулирующие микроРНК передают эту информацию в зародышевые клетки → эпигенетическая модификация устанавливается в зародышевых клетках → гаметы вкладывают эпигенетический фактор в ооцит → фактор распространяется через зиготическое и зародышевое перепрограммирование → фенотипический признак наследуется от неэкспонированного потомства. Модель подтверждается появлением новых фактических данных [Szyf2015b]. Подробный анализ статьи [Szyf2015] сделан в работе {Sharma2015} Abhay Sharma. Transgenerational epigenetic inheritance: resolving uncertainty and evolving biology // Biomolecular concepts 6 (2), 87-103 (2015). DOI 10.1515/bmc-2015-0005 Ниже приведены аннотация м выводы статьи [Sharma2015] Аннотация. О трансгенерационном эпигенетическом наследовании у животных все чаще сообщалось в последние годы. Однако, есть спорные вопросы вокруг этого нетрадиционного способа наследования, особенно у млекопитающих, по нескольким причинам. Во-первых, само существование его было поставлено под сомнение в связи с предполагаемой недостаточностью имеющихся доказательств. Во-вторых, оно потенциально предполагает передачу наследственной информации от сомальной клетки к зародышевой, что противоречит установленному принципу в биологии. В-третьих, по своей природе оно требует выживания через перепрограммирование эпигенетической памяти, что создает еще одну фундаментальную проблему в биологии. В-четвертых, эволюционное значение эпигенетической наследственности также является предметом споров. В данной статье рассматриваются последовательно все эти проблемы на основе последних эмпирических, теоретических и концептуальных достижений. 1) Подробно описываются приведенные основные экспериментальные данные, демонстрирующие возникновение зародышевой эпигенетической наследственности у млекопитающих. 2) Приводятся вновь возникающие доказательства, подтверждающие связи сомальных и зародышевых клеток в трансгенерационном наследования у млекопитающих, и подробно обсуждается роль экзосомы и внеклеточной микроРНК в этой передаче. 3) Выделяется достоверность распространения эпигенетической информации через перепрограммирование. 4) Анализы, поддерживающие эволюционную значимость эпигенетической наследственности, лишь кратко упоминаются. И, наконец, интегративной моделью «эволюционной трансгенерационной системной биологии» предлагается обеспечить основу для будущих достижений в эпигенетической наследственности. Вывод. Все чаще разрушается скептицизм, окружающий существование эпигенетической зародышевой наследования у млекопитающих. Предоставлены убедительные доказательства для возникновения трансгенерационной эпигенетической наследственности у крыс и мышей, которыми проведенные исследования установили возможные последствия и устранили искажающие доводы. Что касается, казалось бы, неправдоподобной коммуникации сомальной и зародышевой клеток, новые данные подтверждают идею, что экзосома и циркулирующие микроРНК могут опосредовать межклеточные связи в эпигенетической наследственности у млекопитающих. Что касается эпигенетической памяти, имеющиеся данные свидетельствуют о том, что информация, содержащаяся в форме метилирования ДНК, модификации гистонов и РНК может передаваться через поколения у животных, включая млекопитающих. Кроме того, накапливается доказательства в пользу роли эпигенетической наследственности в эволюции. Вместе новые достижения в области эпигенетической наследственности расширяют границы современной биологии. Сделанный анализ позволяет вполне аргументированно предполагать, что книга каждой клетки (CellBook) находится не только в этой одной клетке. Это очень важный для модели CellBook для родословной клетки. Время покажет, насколько мы правы. Peter Belobrov 13 Nov 2016 03:16 PIT00351 Перенос РНК к следующему поколению Ещё одна тайна CellBook, которая формирует родословную клеток, видимо, раскрыта! Внеклеточная РНК передаётся от одного поколения к другому в Caenorhabditis elegans. Такие факторы, как питание, стресс, и химические вещества в окружающей среде могут вызвать генетические или эпигенетические изменения, которые передаются от родителей детям, но молекулярные механизмы того, как это может происходить, оставались непостижимы. Один из возможных сценариев (биологическая логика CellBook «клетка – оркестр без дирижёра» включает множество других сценариев, которые рождаются при биологическом взаимодействии без заранее заданной программы!) включает в себя внеклеточные РНК, которые секретируются определенными клетками в организме в ответ на изменения в окружающей среде. Если они усваиваются зародышевыми клетками, то они могут индуцировать изменения ДНК или хроматина, которые сохраняются в следующих поколениях. Начало раскрытия этой загадки было положено в работе {Devanapally2015} Sindhuja Devanapally, Snusha Ravikumar, and Antony M. Jose. Double-stranded RNA made in C. elegans neurons can enter the germline and cause transgenerational gene silencing // Proceedings of the National Academy of Sciences 112 (7), 2133-2138 (2015). doi 10.1073/pnas.1423333112 Cited by 22 times {Marre2016} Julia Marré, Edward C. Traver, and Antony M. Jose. Extracellular RNA is transported from one generation to the next in Caenorhabditis elegans // Proceedings of the National Academy of Sciences (2016), p. 201608959. doi 10.1073/pnas.1608959113 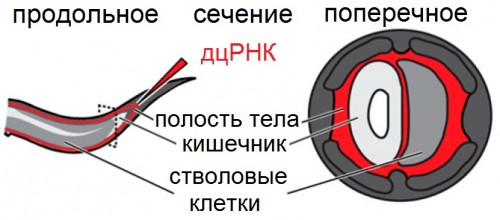 Рис. 3A из [Marre2016]. Схематически показана инъекция дцРНК в полость тела. Внеклеточную дцРНК не надо вводить в цитоплазму любой клетки родителей, чтобы вызвать замалчивание генов (интерференцию) в потомстве. Кроме [Devanapally2015] следуют выделить работы, в которых изучается наследование эпигенетики стволовых клеток, особенно не генетическое наследование и трансгенерационную эпигенетику. Эту биологическую логику рассмотрим в PIT00352 Peter Belobrov 13 Nov 2016 02:14 PIT00350 Геномное редактирование родословной Продолжаю пост PIT00290, в котором обсуждался эксперимент по трассировке родословной целостного организма комбинаторным и кумулятивным геномным редактированием. Вот точная ссылка на статью {McKenna2016} Aaron McKenna, Gregory Findlay, James A. Gagnon, Marshall Horwitz, Alexander Franz F. Schier, and Jay Shendure. Whole organism lineage tracing by combinatorial and cumulative genome editing // Science 353 (6298), 462-aaf7907 (2016). Аннотация DOI 10.1126/science.aaf7907 В отличие от препринта в статье есть подробная аннотация (такие только Science делает) с рис.1. 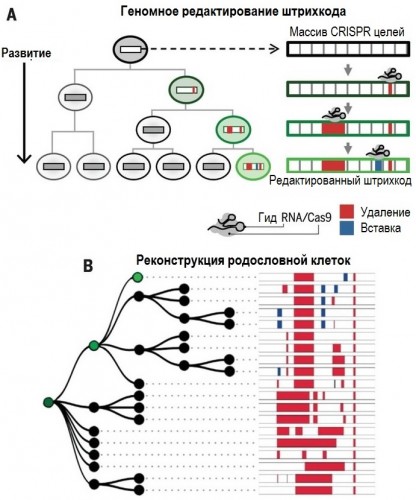 Рис. 1. Целостное восприятие результатов статьи [McKenna2016]. (А) Штрих-код CRISPR/cas9 целевых сайтов постепенно редактируется в течение многих клеточных делений. (B) Отредактированные последовательности штрих-кода связаны друг с другом на основе общих мутаций для восстановления деревьев клеточных родословных. Продвинулся в модели CellBook, обсуждение которой продолжу вскоре. Peter Belobrov 19 Oct 2016 00:00 PIT00339 Центриоли: дочки-матери Этот пост надо читать вместе с PIT00332, чтобы увидеть фундаментальные биологические законы родословной клеток, например, в модели CellBook. Во время мейоза, яйцеклетки теряют центриоли. Исследователи впервые увидели как это происходит.  Первая полная запись В динамике этот процесс показан на видео дочки-матери, где яйцеклетки морской звезды устраняют критические структуры при рождении центриолей в дочерных клетках. Результаты исследования опубликованы в работах . Joana Borrego-Pinto et alt. Distinct mechanisms eliminate mother and daughter centrioles in meiosis of starfish oocytes // The Journal of cell biology 212 (7), 815-827 (2016). DOI 10.1083/jcb.201510083 Ana Pimenta-Marques et al. A mechanism for the elimination of the female gamete centrosome in Drosophila melanogaster // Science (2016): aaf4866. 26 May 2016. DOI 10.1126/science.aaf4866 Matthia A. Karreman et al. Fast and precise targeting of single tumor cells in vivo by multimodal correlative microscopy // J Cell Sci 129 (2), 444-456 (2016). doi 10.1242/jcs.181842 Обсуждение результатов (краткое и понятное) можно найти в статьях . Todd A. Schoborg, and Nasser M. Rusan. Taking Centrioles to the Elimination Round // Developmental cell 38 (1), 10-12 (2016). doi 10.1016/j.devcel.2016.06.027 Marie-Hélène Verlhac. Mother centrioles are kicked out so that starfish zygote can grow // The Journal of cell biology 212 (7), 759-761 (2016). DOI: 10.1083/jcb.201602053 Katie Gerhardt. WORLD OF REPRODUCTIVE BIOLOGY: Vanishing Centrioles in Meiosis // Biology of Reproduction (2016): biolreprod-116. Peter Belobrov 05 Oct 2016 01:22 Мы частично наследуем форму наших ушей, карие глаза, или непокорные волосы нашего отца, но митохондриальная ДНК всегда передается потомству по материнской линии. До сих пор это остается загадкой. Как и почему отцовская митохондрия (митохондрион — гигантская спиральная митохондрия, но почему их на рисунке ниже нарисовано много - не знаю!), находящаяся в сперматозоиде, устраняется, как только эмбрион начинает развиваться. Сперматозоид несёт митохондрион, но его в конечном итоге нет в потомстве. Почему? Новое исследование проливает свет на эту вековую головоломку. Теперь идентифицирован процесс, который устраняет ДНК митохондрии отца во время оплодотворения. {Zhou2016q} Qinghua Zhou et al. Mitochondrial endonuclease G mediates breakdown of paternal mitochondria upon fertilization // Science 353 (6297), 394-399 (2016). DOI 10.1126/science.aaf4777 {Bliek2016} Alexander M. van der Bliek. Demystifying the demise of paternal mitochondrial DNA // Science 353 (6297), 351-352 (2016). Insights Perspectives DOI 10.1126/science.aah4131 {Wang2016} Yang Wang et al. Kinetics and specificity of paternal mitochondrial elimination in Caenorhabditis elegans // Nature Communications 7, 12569 (2016). doi 10.1038/ncomms12569 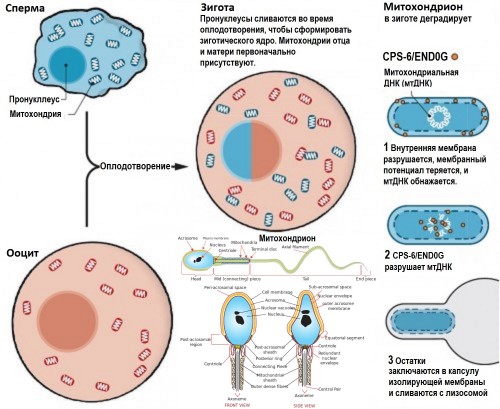 Кнопка самоуничтожения? Отцовская митохондриальная ДНК устраняется во время оплодотворения у разных видов, как это показано на рисунке для червя C. elegans. Модель базового механизма включает нарушение мембраны отцовской митохондрии. Основа рисунка взята из [Bliek2016] с добавлением структуры митохондриона из Wikipedia Spermatozoon. Понимание деталей можно достичь только после чтения статей [Zhou2016q] и [Wang2016]. Занятие не простое, но весьма полезное. Вспоминая деметилирование всей отцовской ДНК, попробуем сказать нечто содержательное о родословной клетки в модели CellBook. Первое, наивное, хотя самое важное то, что стадии деления клеток при сравнении митоза и мейоза описываются канонически и формально. Вот и открывается синаптонемный комплекс, которому сначала приписывается необходимость для генетической рекомбинации. А затем доказывается, что синаптонемный комплекс не является необходимым для генетической рекомбинации. См. подробнее Синаптонемный комплекс и детальнее Synaptonemal complex. Родословная клетки как и модель CellBook требуют более детального рассмотрения, которое мы пока отложим до лучших времён. Peter Belobrov 27 Sep 2016 23:44 PIT00330 Тихоходки - гении выживания Ещё вчера обратил внимание на необычную новость, которую ждал давно в качестве примера горизонтального переноса генов в эукариотических клетках. Дарья Загорская в тексте "Гении выживания – тихоходки подарили человеческим клеткам устойчивость к радиации (21/9/16)" реферировала статью {Hashimoto} Takuma Hashimoto at al (28 авторов). Extremotolerant tardigrade genome and improved radiotolerance of human cultured cells by tardigrade-unique protein // Nature Communications 7, 12808 (2016). Published online 20 September 2016. doi 10.1038/ncomms12808 Abstract. Tardigrades, also known as water bears, are small aquatic animals. Some tardigrade species tolerate almost complete dehydration and exhibit extraordinary tolerance to various physical extremes in the dehydrated state. Here we determine a high-quality genome sequence of Ramazzottius varieornatus, one of the most stress-tolerant tardigrade species. Precise gene repertoire analyses reveal the presence of a small proportion (1.2% or less) of putative foreign genes, loss of gene pathways that promote stress damage, expansion of gene families related to ameliorating damage, and evolution and high expression of novel tardigrade-unique proteins. Minor changes in the gene expression profiles during dehydration and rehydration suggest constitutive expression of tolerance-related genes. Using human cultured cells, we demonstrate that a tardigrade-unique DNA-associating protein suppresses X-ray-induced DNA damage by ∼40% and improves radiotolerance. These findings indicate the relevance of tardigrade-unique proteins to tolerability and tardigrades could be a bountiful source of new protection genes and mechanisms. Вернёмся к тексту новости на русском. Водные медведи или тихоходки (Tardigrade) – это пухлые микроскопические животные размером с песчинку, которые хоть и близки к нематодам и членистоногим по отдельным признакам, но не относятся ни к тем, ни к другим. Всего насчитывается более тысячи видов тихоходок. Большинство видов этих микроскопических полупрозрачных существ размером от 0,5 до 1,5 миллиметра населяют мхи и лишайники. Есть также тихоходки, обитающие в пресных водоёмах и морях. Они обитают в морских и пресных водоёмах, а также на суше во влажных местах, то есть вода для них – это неотъемлемое условие для благополучного роста и размножения. Внешне водные медведи напоминают нечто среднее между гусеницей и голым землекопом и очень гармонично смотрелись бы в каком-нибудь фильме про пришельцев. Но главное, что тихоходки поражают воображение своей способностью к выживанию в экстремальных условиях. Им не страшен ни холод, ни жара, ни огромное давление, ни радиационное излучение. Эти удивительные создания были запущены в открытый космос и некоторые из них после возвращения смогли дать здоровое потомство. В одном из экспериментов тихоходок поместили на 26 часов в ванну с жидким азотом при температуре минус 253˚С. И всё, что потребовалось, чтобы вернуть их к жизни – это несколько капель воды. К слову, самая низкая температура, зафиксированная на Земле, а именно в Антарктиде, – это всего лишь минус 89,2˚С, так что "запас прочности" у тихоходок необычаен. Человеку такие подвиги, мягко говоря, не под силу. Поэтому исследователи изо всех сил пытаются выведать у тихоходок их секреты. При попадании в экстремальную ситуацию водные медведи не пытаются к ней приспособиться, а впадают в анабиоз, то есть у них временно в 10 тысяч раз замедляется обмен веществ. При этом тело тихоходок теряет почти всю жидкость. До сих пор учёные до конца не могут разобраться, как они это делают и какие молекулярные механизмы лежат в основе этих процессов. Большинство специалистов сходятся во мнении, что ключом к успеху тихоходок является их повышенная способность восстанавливать повреждённую ДНК, особенно при выходе из состояния крайнего усыхания. Для того чтобы лучше понять, что происходит в организме водных медведей, исследователи вводят их гены в клеточные культуры млекопитающих, в том числе и человека. Именно так и поступила команда молекулярных биологов из Японии, возглавляемая Такэкадзу Куниэдой (Takekazu Kunieda) из Университета Токио. Для начала учёные секвенировали (определили точное строение) геном тихоходок вида Ramazzottius varieornatus. Представители именно этого вида считаются самыми выносливыми водными медведями. В дальнейшем, экспериментируя с клетками человека и генами тихоходок, исследователи обнаружили, что своеобразным "защитным щитом" для ДНК животных в случае высыхания и воздействия радиации служит белок, известный как Dsup. Белок Dsup (Damage suppressor) достаточно подробно описан в единственной статье [Hashimoto], что достаточно странно. Поэтому, из того ажиотажа, который возник буквально за один день во время написания этого поста, вокруг экстремальных характеристик водного медведя, следует однозначный вывод. Более того, клетки человека, в которых он присутствовал, сами по себе были способны подавлять разрушительное действие радиации на ДНК на 40%. Об этом действительно удивительном открытии сообщается в статье, опубликованной в издании Nature Communications. Специалисты считают, что новые результаты могут иметь важное значение для медицины. Они открывают дорогу к повышению стрессоустойчивости человеческих клеток, что в один прекрасный день может помочь людям, проходящим курс радиационной терапии. Посмотрите видео, где тихоходки - водные медведи показаны живыми. Прекрасное видео, особенно, когда вспоминаешь, что их размер от 0,5 до 1,5 мм! Кроме этого, защитные молекулярные механизмы не повредили бы и тем, кто работает с ядерными установками. А если колонизация Марса всё-таки состоится, то благодаря тихоходкам, вероятно, удастся вырастить на Красной планете первый урожай. Интересно, что данные японских специалистов ставят точку в одном более раннем спорном исследовании, касательно тихоходок. Его авторы секвенировали геном другого вида водных медведей — Ramazzottius hypsibius. Тогда учёные установили, что почти пятая часть генов тихоходок заимствована у других животных и растений, то есть приобретена в процессе так называемого горизонтального переноса. На основе полученных результатов была выдвинута теория о том, что именно высокий процент чужеродных генов и обеспечивает уникальную живучесть тихоходок. Противники выводов нашлись сразу. Они были уверены, что исходные образцы были просто загрязнены. Анализ Куниэды, проведённый с точностью в 100 раз большей, чем спорное исследование, выявил всего лишь 1,2% заимствований. И, поскольку горизонтальный перенос генов оказался ни при чём, молекулярным биологам и другим специалистам придётся ещё немало потрудиться, чтобы разобраться в таком богатом на сюрпризы материале, как геном тихоходки. Посмотрите видео, где за 4 минуты можно увидеть реконструкцию жизни водного медведя. И ещё несколько видео, которые сильнее любой фантастики, хотя показывают жизнь полноценного организма-экстремала размером всего 500 микрон. What Is A Water Bear? = кто такой водный медведь? Water Bear Being A Water Bear = формирование водного медведя. First Animal to Survive in Space = первое животное, выжившее в открытом космосе! How To Find Water Bears = как найти водных медведей? Впрочем, наберите Water Bear в YouTube и размышляйте о том, как клетка становится живым организмом - водным медведем! Peter Belobrov 22 Sep 2016 08:42 PIT00322 Неменделевская родословная В нашей модели CellBook определение наследования как перемешивания клеточных линий можно рассмотреть на интересном примере, взятом из недавней статьи {Besseling2016} Judith Besseling, and Henrik Bringmann. Engineered non-Mendelian inheritance of entire parental genomes in C. elegans // Nature Biotechnology (2016). doi 10.1038/nbt.3643 в которой искусственно создано неменделевское наследование всего материнского или отцовского генома в Caenorhabditis elegans путем изменения структуры митотического веретена во время первого клеточного деления зиготы. Использование зародышевой специфической сверх экспрессии одного белка, сохраняющегося силовым регулятором микротрубочек GPR-1 [см ниже Redemann2011], позволило увеличивать силы, которые тянут полюсами веретена для преобразования одного биполярного митотического шпинделя до двух монопольных шпинделей. Это порождает два клеточных эмбриона, в которых одна клетка содержит только материнские хромосомы, а другая клетка содержит только отцовские хромосомы. В результате этого по мере развития эмбриона, каждая клетка животного, в том числе и зародышевые клетки, содержит генетический материал только одного из родителей в гибридном животном Такую инженерию неменделевского наследования удалось сделать благодаря результатам статьи {Redemann2011} Stefanie Redemann, Siegfried Schloissnig, Susanne Ernst, Andrey Pozniakowsky, Swathi Ayloo, Antony A. Hyman, and Henrik Bringmann. Codon adaptation-based control of protein expression in C. elegans // Nature methods 8 (3), 250-252 (2011). doi 10.1038/nmeth.1565 в которой представлен способ контролировать уровень белка нативной генетической регуляцией в Caenorhabditis elegans с помощью синтетических генов с адаптированными кодонами. Было обнаружено, что сила, действующая на шпинделе в C. elegans эмбрионов, связана с количеством регулятора G-белка GPR-1/2. Кодон-адаптированные версии любого C. elegans гена можно сконструировать с помощью веб-инструмента, или адаптера кодонов C. elegans, созданного авторами статьи [Redemann2011]. Яркое биологическое применение описано в недавней статье Основные моменты статьи Графическое резюме статьи 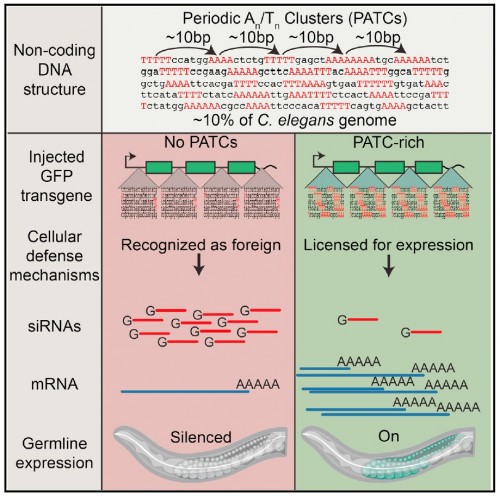 Некодирующие ДНК могут подавить стохастическое замалчивание генов в родословной (Germline) C. elegans. Клетки извлекают пользу от "глушения=замалчивания" чужеродных генетических элементов, но должны одновременно избегать инактивации собственных генов. Хотя модификации хроматина и РНК способствуют поддержанию молчащих состояний, создание молчащих областей неизбежно отражает основную последовательность и/или структуру ДНК. Показано, что распространяющаяся некодирующая особенность ДНК в Caenorhabditis elegans, характеризуется 10 парами оснований периодических Чем отличается менделевское и неменделевское наследования хорошо написал {Perkel2016} Jeffrey M. Perkel. Engineering Sexual Reproduction, Peter Belobrov 08 Sep 2016 10:38 PIT00294 Физика эпигенетики Рекомендую Ruggero Cortini, Maria Barbi, Bertrand R. Caré, Christophe Lavelle, Annick Lesne, Julien Mozziconacci, and Jean-Marc Victor. The physics of epigenetics // Reviews of Modern Physics 88 (2), 025002 (2016). doi 10.1103/RevModPhys.88.025002, Prepint arXiv 1509.04145 (v1 2 Sep; v2 15 Dec 2015; 9 May 2016). http://arxiv.org/abs/1509.04145, 19/4/2020 Cited by 69 Захватывает дух! Но ... для меня это ещё один сигнал, что Принципы биологии клетки ab initio (PIT00174, PIT00220) продолжают развиваться, стремясь к формулировке точных законов фундаментальной биологии ... Основы теории биологических мер. Образ, как и прообраз биологической переменной требует некоторого описания биологического образа аналогичного художественному образу. Аксиоматики био не существует, о чем сказал ещё Гёдель! Ю.М. Лотман ввёл понятие семиосферы по аналогии с биосферой В.И. Вернадского. М. Барбьери создал биосемиотику и с помощью введенных им химических и биологических кодов наравне с культурными кодами PIT00482 сильно продвинулся в понимании биологического смысла аргументов, переменных и значений функций живой природы. Хочу обратить внимание на биосферу как на пространство биологических переменных, функций и значений функций многих биологических переменных. Подчеркиваю, без шуток, что именно это биопространство является правильным образом существования Живой Природы, которая создала и включила в себя живую почву. Конечно же надо согласиться с Владимиром Ивановичем, что ноосфера является пространством существования живого духа. Представление образных значений многих переменных часто разумно представлять в тесном слиянии с количественными характеристиками живых сущностей. Совокупность образных и количественных значений любой биологической переменной без исключений является условной, т.е. переменной, значения которой зависят от состояния всех нижних и верхних уровней. Паттерны изменяются при переходе с уровня на уровень. Создается БиоТеория. Многое прояснилось, пишется работа: «Переменные значения аргументов и функций в логике био». Многомерные пространства значений этих переменных сочетают образные и количественные условные величины. Биоматематика и биологика этих переменных создается для того, чтобы можно было строить точные суждения с биологическими паттернами на различных уровнях. Правила отображения с уровня на уровень надо изучать и раскрывать, т.к. живые объекты содержат программы с созидающими кодами PIT00482, причём биологические механизмы, работающие в рамках первых принципов, достаточно интересные. Многое удаётся понять по результатам организации жизни в переменных, отношениях, событиях, статистиках, условных и совместных мерах, континуумах беспокойных состояний. Самое главное - это выживание созданных вновь артефактов, которое определяется расширениями химических и биологических кодов. Паттерн как волновая функция всей клетки и достаточно изолированных её частей. Передача паттерна преобразованием МакКлинток – органа в геном и обратно - основа теории биологических событий, с помощью которой рассматриваются самые главные состояния, преобразования их, измерение свойств с точки зрения биологического действия. Понятие эквивалентности неприменимо для разных типов переменных. Так любые количественные значения невозможно сопоставить с образными значениями картин, стихов, музыки, танцев и, вообще, любой экспрессии. Тип отображения «экспрессия» требует тщательного рассмотрения. Кроме эквивалентности условность переменных высокой размерности потребуется особые усилий для согласования неколичественных характеристик роста и развития ... Обновление 19/4/2020 «Физическая структура и структура данных трехмерного генома» Kai Huang, Yue Li, Anne R. Shim, Ranya KA Virk, Vasundhara Agrawal, Adam Eshein, Rikkert J. Nap, Luay M. Almassalha, Vadim Backman, and Igal Szleifer. Physical and data structure of 3D genome // Science Advances 6 (2), eaay4055 (2020). doi 10.1126/sciadv.aay4055. «ChIPSummitDB: база данных сайтов связывания транскрипционных факторов на основе ChIP-seq и топологическое расположение белков, связанных с ними» Erik Czipa, Mátyás Schiller, Tibor Nagy, Levente Kontra, László Steiner, Júlia Koller, Orsolya Pálné-Szén, and Endre Barta. ChIPSummitDB: A ChIP-seq-based database of human transcription factor binding sites and the topological arrangements of the proteins bound to them // Database 2020, baz141 (2020). doi 10.1093/database/baz141 «Структура хроматина изменяется во время различных процессов с точки зрения последовательности ДНК» Hui Quan, Ying Yang, Sirui Liu, Hao Tian, Yue Xue, and Yi Qin Gao. Chromatin structure changes during various processes from a DNA sequence view // Current opinion in structural biology 62, 1-8 (2020). doi 10.1016/j.sbi.2019.10.010 «Различение клеточного фенотипа с использованием клеточного эпигенотипа» Thomas P. Wytock, and Adilson E. Motter. Distinguishing cell phenotype using cell epigenotype // Science Advances 6 (12), eaax7798 (2020). doi 10.1126/sciadv.aax7798 «Количественная оценка эпигенетической стабильности с минимальными путями действия» Amogh Sood, and Bin Zhang. Quantifying Epigenetic Stability with Minimum Action Paths // Preprint bioRxiv 20 Feb (2020). doi 10.1101/2020.02.25.964726 Peter Belobrov 09 Jul 2016 05:50 PIT00293 Как клетка организует органеллы Статью надо прочесть (она в открытом доступе) и посмотреть прилагаемых 10 видео. Результат превзойдёт все ваши ожидания. {Lin2016} Congping Lin, Martin Schuster, Sofia Cunha Guimaraes, Peter Ashwin, Michael Schrader, Jeremy Metz, Christian Hacker, Sarah Jane Gurr, and Gero Steinberg. Active diffusion and microtubule-based transport oppose myosin forces to position organelles in cells // Nature communications 7, 11814 (2016). doi 10.1038/ncomms11814 Добавление 11/10/2016 Журнал BioTechniques снова повторил сообщение, опубликованное 15/6/2016. Редакция понимает важность нахождения ответа на вопрос: "Как клетка организует органеллы?" Понятно моё решение посмотреть на то, как реагировала научная общественность на [Lin2016]. Пока только пара цитирований, но каких! Микротрубочки достаточно изученный объект, но в статье [Lin2016] был затронут вопрос о дальнодействующих биологических взаимодействиях. Поэтому в одной статье [Salogiannis2016] (In Press, Available online 21 September 2016) рассмотрены канонический и неканонической режим (автостоп или Hitchhiking) поведения микротрубочек, связанный с межклеточным транспортом, а именно, движения на больших расстояниях взаимодействующих органелл, везикул и макромолекулярных комплексов микротрубочек на основе транспорта, что имеет решающее значение для роста клеток и выживания. 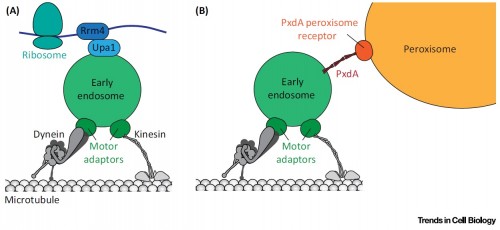 Механизмы перемещения автостопом [Salogiannis2016]. В текстовом варианте этой статьи есть полезный для понимания глоссарий, которого нет в файле pdf. Да и будущие направления работы сформулированы нешуточные. Например, Каково биологическое значение автостопа? Какие сигналы запуска и останови переноса автостопом? Сохраняется ли автостоп в клетках млекопитающих? Не слабые вопросы! Вот где можно помечтать про изучение автостопа в микрофлюидном чипе! {Salogiannis2016} John Salogiannis, and Samara L. Reck-Peterson. Hitchhiking: A Non-Canonical Mode of Microtubule-Based Transport // Trends in Cell Biology (2016). Available online 21 September 2016. In Press doi 10.1016/j.tcb.2016.09.005 В другой работе [Matei2016] (декабрь 2016 !) рассмотрена пузырчатая головня Ustilago maydis, которая является известной моделной системой для биотрофных грибковых патогенов растений. Гриб имеет диморфной жизненный цикл с сапрофитный фазы и дрожжеподобным переходом на нитчатый патогенный рост. Поэтому из-за такой сильной дифференцировки клеточная биология кукурузной головни Ustilago maydis рассмотрена в качестве модели биотрофных взаимодействий. {Matei2016} Alexandra Matei, and Gunther Doehlemann. Cell biology of corn smut disease — Ustilago maydis as a model for biotrophic interactions // Current Opinion in Microbiology 34, 60-66 (2016). December 2016 ! doi 10.1016/j.mib.2016.07.020 Peter Belobrov 16 Jun 2016 02:06 PIT00290 CellBook - эксперимент Мне трудно даже было представить, что экспериментальное исследование родословной клеток начнётся так быстро. Поэтому, когда на сайте vesti.ru встретил новость На "пленку" ДНК записали историю жизни всех клеток, то был в шоке. Стал проверять - мало ли что напишут журналисты, у которых генетический код меняется при каждом делении (к сожалению, некоторые студенты так думают, принимая геном за генетический код). Но когда прочитал об использовании биоштрихкода при изучении родословной клеток, то понял, что "лёд тронулся, господа присяжные заседатели". Замечу здесь, что на этом сайте понятие родословной клетки используется давно, т.к. с моей точки зрение понятие клеточной линии (lineage) не позволяет грамотно описать биологию развития организмов, когда сетевой и стволовой принципы при симметричном и ассиметричном делении перемешиваются. Надо строить точную модель родословной! Вот препринт этой статьи Удивительно, что c этой новостью русские вести опередили BioTechniques на 3 дня, хотя статья Jeffrey M. Perkel. Tracing Cell Lineages with CRISPR/Cas // BioTechniques 3/6/2016 более сдержанная. Модель CellBook надо доводить "до ума"! Peter Belobrov 31 May 2016 13:14 PIT00279 Биологическое разделение фаз При подготовке этого поста пришла мысль о возможности чтения и записи клеточной родословной путём биологического разделения фаз. Насколько это "наследуемо" в CellBook - вопрос открытый пока, но мы достаточно близки к его точной постановке, особенно, если включим в рассмотрение разрыв сплошности жидкой фазы твёрдой частицей (липидным рафтом = плотиком) или газовой прослойкой. От лирического начала биологического разделения фаз или P-гранул PIT00003 организация РНК в новом свете PIT00207 (добавил рисунок и дал ссылку открытого доступа) продолжает привлекать внимание. В перекрестии ссылок была найдена функциональная организация высшего порядка через разделение фаз в биологии, которая рассмотрена в статье Diana M. Mitrea, Richard W. Kriwacki. Phase separation in biology; functional organization of a higher order // Cell Communication and Signaling 14 (1), 1 (2016). DOI 10.1186/s12964-015-0125-7 (Прекрасный обзор в открытом доступе. Рекомендую прочесть!) Abstract. Inside eukaryotic cells, macromolecules are partitioned into membrane-bounded compartments and, within these, some are further organized into non-membrane-bounded structures termed membrane-less organelles. The latter structures are comprised of heterogeneous mixtures of proteins and nucleic acids and assemble through a phase separation phenomenon similar to polymer condensation. Membrane-less organelles are dynamic structures maintained through multivalent interactions that mediate diverse biological processes, many involved in RNA metabolism. They rapidly exchange components with the cellular milieu and their properties are readily altered in response to environmental cues, often implicating membrane-less organelles in responses to stress signaling. In this review, we discuss: (1) the functional roles of membrane-less organelles, (2) unifying structural and mechanistic principles that underlie their assembly and disassembly, and (3) established and emerging methods used in structural investigations of membrane-less organelles. 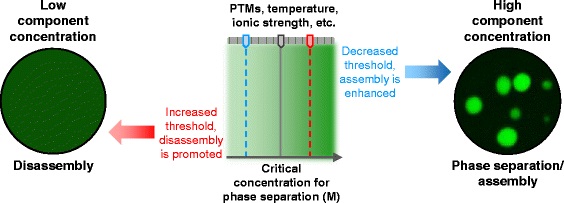 Рис. 1. Macromolecular condensation mediates the formation of membrane-less organelles. Membrane-less organelles are dynamic structures formed via a polymer-condensation-like, concentration-dependent phase separation mechanism. The critical concentration threshold (grey line) for phase separation can be tuned within a range of concentrations (shaded green box) through physico-chemical alterations to the system (i.e., posttranslational modifications to domains and/or motifs that alter the affinity of their interactions, changes in temperature, altered ionic strength, etc.). These changes can drive phase separation and assembly of membrane-less organelles, or their disassembly. 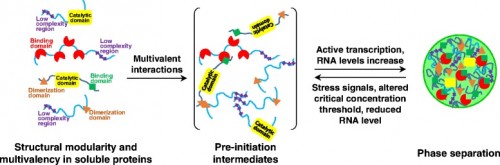 Рис. 2. Molecular basis for membrane-less organelles assembly. The proteins enriched within the matrices of membrane-less organelles commonly exhibit multiple modules that create multivalency, including folded binding domains (red) and low complexity regions (purple). Valency is often amplified by domains that enable homo-, or hetero-oligomerization (orange). Interactions between proteins containing different combinations of these interaction modules provide a framework for building a heterogeneous, infinitely expandable network within membrane-less organelles. Formation of this type of network drives phase separation when the critical concentration threshold is reached. For many of the examples discussed herein, active RNA transcription is needed for membrane-less organelle assembly. We hypothesize that expression of RNA in excess of a critical concentration threshold is needed to nucleate interactions with specific, multi-modular proteins, and for nucleating formation of membrane-less organelles. Stress signals can alter the multivalent interactions that drive phase separation and lead to partial or complete disassembly of the organelle. В этот обзоре есть достаточно полный список немембранных (membrane-less) органелл клетки, в который включены белки и РНК 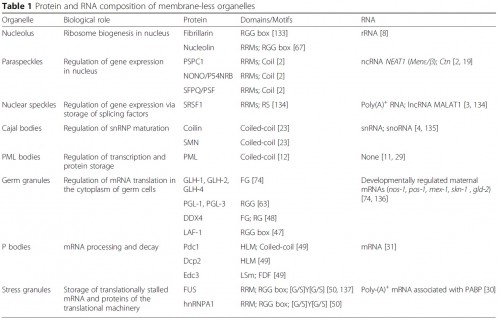 Peter Belobrov 23 Feb 2016 13:12 PIT00266 Болезни симбиозов Это очень важный пост! Но так как сейчас почему-то никто не пишет на МОЛПИТе по какой-то странной причине, да и кажется не читает, то придётся исключить из проблемы болезни и иммунитет растительных симбиозов, которому был посвящён пост PIT00253. Давайте рассмотрим опухоли. Вот-вот! Именно, неуправляемый рост клеток, которым не почём все биологические сигналы в симбиозе, говорящие клеткам что-то вроде "хватит, ребята, резвиться, веселиться и размножаться, а то вы гнев медиков на себя навлечёте, и они вас всех и химиотерапией и гамма лучами ... и ...видите какая армия их в раковых институтах и какое число новых методов ...". Позволю себе здесь высказать предположение, основная мысль которого родилась лет ~ 50 назад в беседе с моим другом Володей Галкиным. Как физики мы не имели никакого понятия о симбиозах и о биологических взаимодействиях и о биологической иерархии ... да мало ли чего за 50 лет в биологии было открыто. Так вот. Родилась мысль, что кроме рака человечество ожидают ещё более коварные болезни. Да, тогда ещё не было спида, вирусов Эболы и Зика да и пандемий гриппа таких жестоких не было 50 лет назад. ВОЗ предупредил сегодня (28/2/2016), что скоро почти все Западное полушарие охватит эпидемия лихорадки Зика. http://www.vesti.ru/doc.html?id=2713349&tid=109445 Рассмотрим вызовы эпигенетическим правилам со стороны онкогенов. Прояснения в известном семействе генов-супрессоров опухолей может изменить наше понимание регуляции транскрипции и ключевой роли TET белков в поддержании целостности генома. Результаты острой потери TET-функции в агрессивном миелоидном раке мышей рассмотрены в статье {An2015} Jungeun An, Edahí González-Avalos, Ashu Chawla, Mira Jeong, Isaac F. López-Moyado, Wei Li, Margaret A. Goodell, Lukas Chavez, Myunggon Ko, and Anjana Rao. Acute loss of TET function results in aggressive myeloid cancer in mice // Nature communications 6, 10071 (2015). doi 10.1038/ncomms10071 в открытом доступе 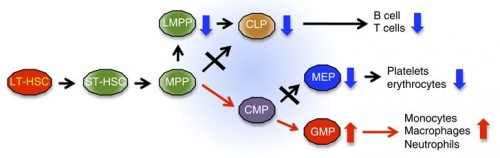 Рис 3h {An2015} Схематическое представление изменений развития кроветворения в Tet2/3 DKO мышах. Сокращения - LT-HSC & ST-HSC, long-term & short-term haematopoietic stem cells; MPP, multipotent progenitors; LMPP, lymphoid-primed multipotent progenitor cells; Миелоидные предшественники (CMP, common myeloid progenitors; GMP, granulocyte–monocyte progenitors; MEP, megakaryocyte-erythroid progenitors; CLP, common lymphoid progenitors); platelets = тромбоциты; миелоидная линия клеток крови характеризуется тем, что быстро размножаются изменённые клетки белой крови. Накапливаясь в костном мозге, они подавляют рост нормальных клеток крови, что приводит к снижению количества эритроцитов, тромбоцитов, и нормальных лейкоцитов. Теперь рисунок полностью понятен! Для справки пара уточнений. ТЕТ-семейство диоксигеназ окисляют 5-метилцитозин (5mC) в ДНК и оказывается супрессором (геном, подавляющий экспрессию других генов) опухоли во многих видах рака. Даже при отсутствии мутаций в TET кодирующей области, потеря ТЕТ-функции сильно связана с раком. Ферменты ТЕТ(ten-eleven translocation = 10-11 транслокация)-семейства являются диоксигеназами, которые преобразуют 5-метилцитозин (5mC) до 5-гидроксиметилцитозин (5hmC) и другие продукты окисления 5-формилметилцитозина (5FC) и 5-карбоксилцитозина (5caC). Вместе эти окисленные метилцитозины (окси-MC) содействуют деметилированию ДНК, а также функционируют в качестве эпигенетических факторов. Потеря их функций и мутации в TET2 связаны с различными миелоидными и лимфоидными злокачественных новообразованиями в организме человека. Прогениторные клетки (клетки-предшественники, но не стволовые клетки!) — стволовые клетки, детерминированные на дифференцировку в определённый тип клеток. Это клетки, которые, в отличие от плюрипотентных стволовых клеток уже имеют стойкие биомаркеры, которые позволяют отличить их и их потомство от клеток других типов. Кроме того их способность к пролиферации значительно ниже чем у плюрипотентных стволовых клеток. Прогениторные клетки выполняют роль стволовых клеток взрослого организма занятых пополнением пула специализированных клеток организма. Различают мультипотентные и унипотентные прогениторные клетки. Иногда термин прогениторная клетка заменяют на термин клетка-предшественница или бластная клетка. Однако следует отметить, что в эмбриологии и цитологии клетками-предшественницами обычно называют группу клеток которые позднее дифференцируются в конкретный орган (PIT: взгляд на клетку со стороны ткани и со стороны прокариоты на эукариоту - имеет отношение к родословной клеток и CellBook). Может и не зря потратил пару часов на этот пост? Кто знает, "как слово наше отзовётся?" Peter Belobrov 28 Jan 2016 03:50 PIT00259 Взаимодействие генов в геноме В России всё меньше лабораторий мирового уровня. Сергей Владимирович Разин держит мировой уровень! Надо мной уже посмеиваются по поводу многолетней постоянной рекомендации студентам прочитать книгу С. В. Разин, А. А. Быстрицкий. Новая статья из лаборатории С.В. Разина опубликованная в журнале Cold Spring Harbor Lab Sergey V. Ulianov, Ekaterina E. Khrameeva, Alexey A. Gavrilov, Ilya M. Flyamer, Pavel Kos, Elena A. Mikhaleva, Aleksey A. Penin et al. Active chromatin and transcription play a key role in chromosome partitioning into topologically associating domains // Genome research (2015). doi 10.1101/gr.196006.115 говорит о том, как происходит взаимодействие генов, когда активный хроматин и транскрипции играют ключевую роль в разделении хромосом на топологически связанные домены. 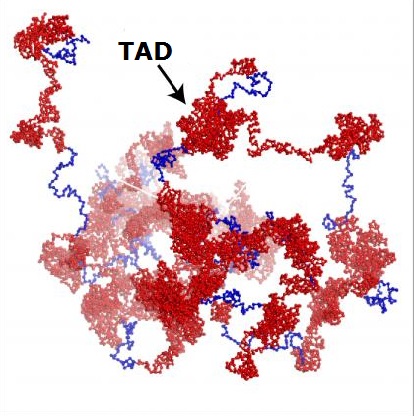 TAD - топологически связанный домен (topologically associating domain). 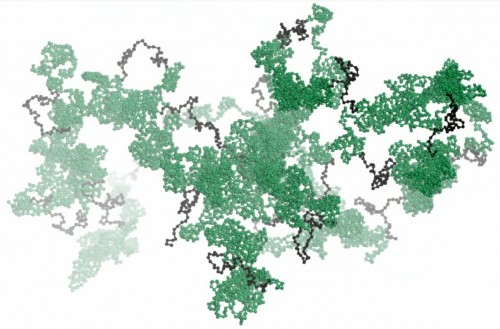 Кадр из видео фильма к статье. В основе предположение, что TADs может самоорганизоваться в зависимости от того, содержит ли участок ДНК активный или неактивный хроматин. Вот эпиграф свежей статьи: От стай птиц до кишащих молекул физики пытаются понять "активное вещество", глядя на фундаментальную теорию живого мира. (From flocking birds to swarming molecules, physicists are seeking to understand 'active matter' — and looking for a fundamental theory of the living world.) Gabriel Popkin. The physics of life // Nature 529, 16–18 (07 January 2016). Статья в открытом доступе, очень интересны комментарии к ней Peter Belobrov 14 Jan 2016 06:00 PIT00256 Коннектом или схема связей мозга? Можно долго подшучивать над коннектомикой (connectomics), но прогресс в её развитии настолько сильный, что даже использование англоязычных калек не облегчает понимания сути дела. Этим постом хочу сказать, что уже доказано, что схема связей мозга настолько разнообразна и нетривиальна, что старые модели нейронных сетей не просто примитивны, но и никуда не годятся! Желание глубже разобраться в новых типах нейронов PIT00254 связано с необходимостью знать целый ряд новых понятий. Внутриклеточный транспорт является направленным; транспорт от эндоплазматического ретикулума через аппарат Гольджи к лизосомам, эндосомам или клеточной поверхности называется антероградным; транспорт в обратном направлении – ретроградным. Кроме диффузной магнито-резонансной томографии (дМРТ) применяется технология Brainbow: Брэйнбоу – это метод нейровизуализации, в основе которого лежит использование флуоресцентных белков. Будучи внедрённым в геном животного, зелёный флуоресцентный белок и его генетически модифицированные варианты окрашивают нервные клетки в разные цвета (в общей сложности до 100 разных оттенков), что позволяет значительно более точно локализовать архитектуру нейронных связей отдельных клеток. Данный метод позволяет картографировать одновременно до 100 нервных клеток. Названием метода является сочетание английских слов brain (мозг) и rainbow (радуга). Несколько аксонов сетчатки (> 10 !) сходятся на переключающих клетках таламуса взрослых мышей. Это доказано в работе Аннотация статьи {Ohno2016}, которая появилась в сети 15/12/15. Продвижением микроскопических технологий создан значительный прогресс в нашем понимании мозга. Недавние усилия по выяснению полной карты электрических связей мозга называемой «коннектом» с помощью различных форм технологии обработки изображений, в том числе световой и электронной микроскопии, начали давать существенный вклад на нескольких организмах. Вклад был бы невозможен без недавних инноваций при получении и анализе больших массивов коннектомных данных. Текущие данные показали, сложные сети с однонаправленными и взаимными связями головного мозга схем на макро- и световой микроскопии («мезоскопических») уровнях, и невообразимой сложности синаптических связей между аксонов и дендритов на электронно-микроскопических («микроскопическом») уровне. В то же время, данные подчеркнул необходимость добиться существенного прогресса в методологии проведения коннектомных исследований, в том числе эффективной обработки и автоматизированного анализа полученных данных. Дальнейшему пониманию структурного и функционального коннектома, по-видимому, способствует комбинация различных методов визуализации. Такие многопрофильные подходы дают нам ключ к решению, может ли полная коннектомика выяснить фундаментальные механизмы обработки основных и высших функций человеческого мозга. Ключевые слова: коннектом, синапс, схема мозга, тректография (trajectography) {Ohno2016} Nobuhiko Ohno, Mitsuhiko Katoh, Yurika Saitoh, and Sei Saitoh. Recent advancement in the challenges to connectomics // Microscopy (2015): dfv371. Advance Access article. Perhaps it will Microscopy 65 (1), dfv371, p.?-? (2016). doi 10.1093/jmicro/dfv371 Для тех, кто будет читать статью, привожу перевод подписей к рисункам (для размещения рисунков и статьи здесь нет разрешения). Рис. 1. Несколько подходов, используемых в исследованиях по коннектомике в разных масштабах. Коннектомные исследования выполняются на макроскопическом (~10-1-10-3 м, для целого мозга траекторий аксонов пучков), мезоскопических (~10-2-10-7 м, для клеточной морфологии и нервных прогнозов) и микроскопические (~10-4-10-9 м, для субклеточных структур, включая синаптических связей) уровней с различными разрешениями (а). Макроскопические, мезоскопические и микроскопические исследования в основном используют дМТР (b), световой микроскопии с флуоресцентным маркировки (c) и 3D реконструкции следующем последовательного приобретения изображения с помощью электронного микроскопа (d), соответственно. Изображения взяты из Шибата и др. [13] (b), Мияваки [47] (c) и Takemura [59] (d). Рис. 2. Различные методы, используемые для маркировки мезоскопических анализа коннектома головного мозговой. В подходе Брэйнбоу (a), генетическая рекомбинация посредничестве системы Cre-LoxP приводит к стохастической маркировки нейронов с комбинациями различных флуоресцентных белков, что позволяет различать траектории отдельных нейронов [36]. С другой стороны, в последнее время исследования использовали коннектомные маркировки определенных нейронов с антероградной (b) и антероградной и ретроградной комбинаций (c) отслеживание методы через инъекции в определенных участках головного мозга [40,45]. FG: fluorogold (золото, меченное флуоресц.меткой); BDA: биотинилирован¬ный декстран амин; PHAL: лейкоагглютинин обыкновенной фасоли; CTb: субъединица b холерного токсина. Разрез мозга мыши с инжектированных антероградным индикатором и контрастирующим 4',6-диамидино-2-фенилиндол (DAPI) (d) показывает, что экспрессия (проявление) EGFP (d, f, g) после впрыскивания аденоассоциированного вируса и обычной антероградной метки BDA (d, e, g) локализуется в нейронах вблизи места инъекции. Область отмечены (d) увеличивается в (e)–(g) и (g) получают путем объединения (e) (показывая BDA) и (f) (показывая EGFP). Полоска: 1000 мкм. Вставки (d)–(g) адаптированы из Oh и др. [40] с разрешения. Peter Belobrov 09 Jan 2016 16:10 PIT00254 Новые типы нейронов Интересное открытие сделано совсем недавно. Традиционно считалось, что неокортекс содержит два основных типа нейронов: пирамидальные нейроны (~80 % нейронов неокортекса) и вставочные нейроны (~20 % нейронов неокортекса). Новая кора (синонимы: неокортекс, изокортекс; лат. neocortex) — новые области коры головного мозга, которые у низших млекопитающих только намечены, а у человека составляют основную часть коры. Новая кора располагается в верхнем слое полушарий мозга, имеет толщину 2—4 миллиметра и отвечает за высшие нервные функции — сенсорное восприятие, выполнение моторных команд, осознанное мышление и, у людей, речь. Используя новый метод изучения принципов подключения между морфологически определенных типов клеток в неокортексе, исследователи под руководством X Jiang и A Tolias обнаружили новые виды нейронов в головном мозге. Xiaolong Jiang, Shan Shen, Cathryn R. Cadwell, Philipp Berens, Fabian Sinz, Alexander S. Ecker, Saumil Patel, and Andreas S. Tolias. Principles of connectivity among morphologically defined cell types in adult neocortex // Science 350 (6264), aac9462 (2015). https://goo.gl/2zuazo 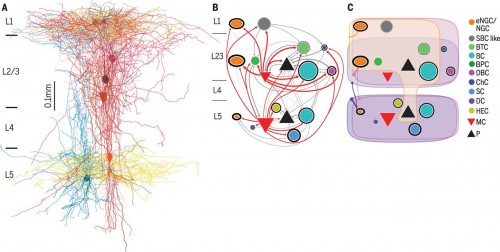 Связь между морфологически определенными типами клеток в неокортексе. (A) Одновременные восьмикратные записи активностей целых клеток для изучения связи с последующей морфологической реконструкцией. (B) Синаптические связи между морфологически различных типов нейронов, в том числе пирамидальных (P) нейронов. (C) Соединения нейроглиальных клеток (NGCs) с другими типами клеток. Эти связи, как полагают, несинаптические и опосредованы объемной передачей (?биологической и/или химической связью?). Martinotti cell, MC (клетки Маринотти); neurogliaform cell, NGC (нейроглиаформные клетки); basket cell, BC (корзиночные клетки); single-bouquet cell-like cell, SBC-like (однобукетно подобные клетки); bitufted cell, BTC (двухохолковые клетки); bipolar cell, BPC (биполярные клетки); double-bouquet cell, DBC (двухбукетные клетки); chandelier cell, ChC (люстровые клетки); shrub cell, SC (кустарниковые клетки); horizontally elongated cell, HEC (горизонтально вытянутые клетки); deep-projecting cell, DC (глубоко выступающие клетки). Эта же команда продолжила электрофизиологическое, транскриптомное и морфологическое профилирование отдельных нейронов с помощью совместного изучения физиологии нейронов и анализа РНК последовательностей (Patch-SEQ метод). Cathryn R. Cadwell, Athanasia Palasantza, Xiaolong Jiang, Philipp Berens, Qiaolin Deng, Marlene Yilmaz, Jacob Reimer et al. Electrophysiological, transcriptomic and morphologic profiling of single neurons using Patch-seq // Nature biotechnology (2015). Accepted 02 Dec 2015; Published online 21 Dec 2015. Фрагмент аннотации. Несмотря на важность неокортекса млекопитающих в сложных познавательных процессах, нам по-прежнему не хватает всестороннего описание его клеточных компонентов. Для улучшения классификации типов нейронов клеток и функциональной характеристике отдельных нейронов, представлен Patch-SEQ, метод, который сочетает в себе записи лоскутным электрофизиологическим зажимом целых клеток, одноклеточные РНК-последовательности и морфологическое описание. Использованием этого подхода созданы электрофизиологические и молекулярные профили 58 клеток неокортекса и показано, что паттерны экспрессии генов могут быть использованы для вывода морфологических и физиологических свойств, таких как разветвление аксонов и амплитуд потенциала действия отдельных нейронов. Эти результаты проливают свет на молекулярные основы разнообразия нейронов и предполагают, что Patch-SEQ метод может способствовать классификации типов клеток в нервной системе. Прекрасный Рождественский подарок всем нам! Peter Belobrov 06 Jan 2016 04:24 PIT00232 Геометрия создает зародышевые листки Думаю, эта уже классическая работа (прочтите сами краткое изложение статьи http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4341966/ Aryeh Warmflash, Benoit Sorre, Fred Etoc, Eric D. Siggia, Ali H. Brivanlou. A method to recapitulate early embryonic spatial patterning in human embryonic stem cells // Nature methods 11 (8), 847–854 (2014). в которой авторы считают, что геометрический контроль клеточной культуры должен стать стандартной практикой для эмбриональной специализации стволовых клеток. Микропаттерновая дифференцировка может быть использована для сравнения межвидовых, а также сравнение между индуцированными плюрипотентными стволовыми клетками человека и ЭСК других видов в аналогичных условиях анализа. 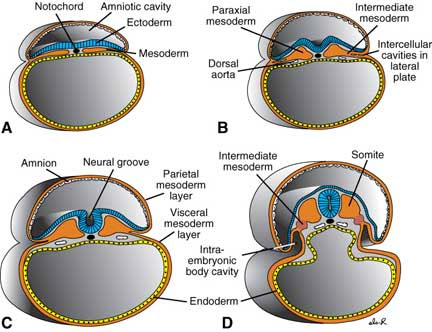 Схема гаструляции у млекопитающих Среди 30 ссылок на эту статью есть статья о достижениях в микрофлюидных платформах, которые использованы для анализа и регулирования человеческих плюрипотентных стволовых клеток Tongcheng Qian, Eric V. Shusta, Sean P. Palecek. Advances in microfluidic platforms for analyzing and regulating human pluripotent stem cells // Current opinion in genetics & development 34, 54-60 (2015). В этой статье концепция микрофизиологических систем немедленно привлекла внимание. Peter Belobrov 13 Oct 2015 05:24 PIT00222 Капельный штрих-код Появился новый тип биоштрихкода - капельный Allon M. Klein, Linas Mazutis, Ilke Akartuna, Naren Tallapragada, Adrian Veres, Victor Li, Leonid Peshkin, David A. Weitz, and Marc W. Kirschner. Droplet Barcoding for Single-Cell Transcriptomics Applied to Embryonic Stem Cells // Cell 161 (5), 1187-1201 (2015). doi 10.1016/j.cell.2015.04.044 Этот эксперимент приближает к пониманию новой парадигмы экспрессии! 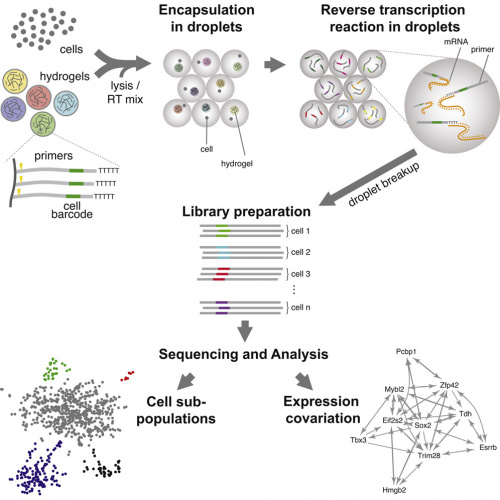 Графическая аннотация платформы для ДНК-штрихкодирования тысяч клеток. Peter Belobrov 27 Aug 2015 01:46 PIT00220 Принципы биологии клетки ab initio Вот и разродился PIT текстом пленарного доклада в Ростове Актуальные проблемы биологии, нанотехнологий и медицины - МГУ: 2015 П.И. Белобров Фундаментальная биология как точная наука накопила гигантский объём знаний, требующих новых подходов к пониманию основных биологических законов. В докладе сделан анализ главных принципов биологии клетки для физической теории живых систем. Принципы клеточной биологии, развивающие традиционный подход [1], следующие. Из принципа постоянного биологического беспокойства следует формула: «действие – это закон жизни». Неравенство действия и противодействия есть движущая сила развития. Отсутствие точек покоя рассмотрено на примере клеточного цикла [2], основных эпигенетических модификаций и родословной стволовых клеток [3]. Биологические взаимодействия имеют многомерную природу, которая гораздо шире комбинаторики белок-белковых сетей (interactome) и требует использования мер Хаусдорфа, описывающих размерность вместе с процессами образования и разрыва связей, изменяющих размерность. Структура и функции клеток обеспечивают гармонию непрерывности «струны жизни» и дискретности организмов, формируя канторовы континуумы состояний клеток и тканей. Принцип биологического разнообразия поддерживает целостность живых организмов, обеспечивая непрерывность жизни в целом. Принцип созидания выражается формулой: «способность создавать и есть жизнь», который является одним из главных свойств живых клеток [4]. Биологические сигналы принципиально зависят от приёмников, преобразующих полученные сигналы и раскрывающих их биологический смысл. Этот принцип рассмотрен на трёх примерах: везикулы межклеточного обмена, сигнальные цепи феромонов насекомых и преобразователи генетических сигналов [5, 6]. Принцип иерархии, организации и сложности генов белков и различных РНК лежит в основе биологии клетки и активно развивается в свете новой парадигмы экспрессии. Родословная клетки определяется принципом перемешивания клеточных линий при наследовании, когда экспрессия генетического материала реализуется ажурной сетью взаимодействия различных РНК и белков в цитоплазме и в хромосомных территориях [7]. Принцип глубокой гармонии индивидуальности и целостности рассмотрен на примерах биологических симбиозов, где согласованы врождённые иммунитеты бактерий, грибов, растений и животных. Эти представления развивались от открытия фагоцитов И.И. Мечниковым через иммунитет растений Н.И. Вавилова до современного понимания [8]. Принцип измерения клетки клеткой [9] и биологические числа [10] позволили определить биологические меры и калибровки, разработка которых начата автором доклада. Понимание биологической информации в клеточных процессах началось с теории Колмогорова [11], следуя которой можно определить сложность как длину алгоритма, преобразующего одну клетку в другую. Соответствующая иерархия кодов и уровней организации расширяет понятие обычной энтропии, хотя начатое в [11] развитие условной энтропии меры по мере ещё далеко не закончено в биологии клетки [5-8]. Много тайн раскрыла теория Е.В. Кунина, в которой показано, что логика случая лежит в основе последовательности биологических событий [12], где точное понимание рождения слабой доминанты требует глубоких исследований. В докладе предложены математические методы описания, а в трудных случаях введены необходимые условия для развития биологической математики (biomathic), где нет аксиом, т.к. биология клетки не является формализуемой наукой. Предложенные принципы – основа клеточной родословной, записанной в молекулярной книге клетки (Cell Book), которая читается самой клеткой и перезаписывается при каждом клеточном делении [2]. Фундаментальная биология точна по биологическим мерам, многие из которых лишь интуитивные, хотя позволяют создавать молекулярные устройства, использующие принципы биологии клетки ab initio. Благодарю Л.И. Патрушева, А.Г. Малыгина и Н.В. Пашенову за обсуждения основной идеи и полезные критические замечания. Работа выполнена за счет средств государственного задания на проведение фундаментальных исследований РАН (проект № 01201351504) и за счет средств гранта Российского научного фонда (проект №15-19-10041). Литература. Peter Belobrov 25 Aug 2015 14:14 PIT00207 Организация РНК в новом свете Многотомная энциклопедия eLife (у нас есть электронное издание 2011 г. ~ из 27 000 страниц) сейчас стала открытым журналом eLife v1-v5 (2012-2016) http://www.ncbi.nlm.nih.gov/pmc/journals/1894/ Новая статья в журнале eLife в некотором смысле сюрприз. Метод решеточной светлопольной микроскопии позволил увидеть набор ленточных белков обернутый вокруг одного типа РНК-гранул. Это, пожалуй, самая интересная новость после длительного затишья с P-гранулами на МОЛПИТ. Jennifer T. Wang, Jarrett Smith, Bi-Chang Chen, Helen Schmidt, Dominique Rasoloson, Alexandre Paix, Bramwell G. Lambrus, Deepika Calidas, Eric Betzig, and Geraldine Seydoux. Regulation of RNA granule dynamics by phosphorylation of serine-rich, intrinsically disordered proteins in C. elegans // eLife 3, e04591 (2014). (Добавление 23/2/2016). Доступ к статье открытый. Можно прочитать всю статью, рассмотреть все рисунки в самом высоком разрешении и посмотреть интересные видео: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4296509/. После этого идея биологического разделения фаз придёт к Вам сама собой. 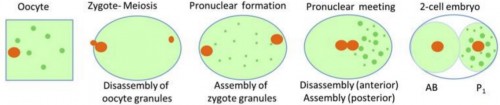 Рис.1а. Динамика Р-гранул во время перехода от яйцеклетки к эмбриону. Зеленые точки - P-гранулы, бледно-зеленый цвет представляет компоненты P-гранул, которые диффундируют в цитоплазму. Оранжевый цвет - пронуклеусы. P1 - зародышевый бластомер. Peter Belobrov 28 May 2015 12:08 Антон поднял вопрос об алгоритме и мы мгновенно переключились на клеточные алгоритмы, которые очевидно, не цифровые, но какие? Было это, пожалуй, в четверг 2 апреля. Аналоговые "вычисления", измерения без цифр, аналоговые биологические меры ... новый мир клеточных книг, т.е. CellBook или родословной клеток. И вот http://www.computer-museum.ru/frgnhist/mathem_21.htm текст А.С. Нариньяни "Математика 21 – радикальная смена парадигмы: Модель, а не Алгоритм". Прочтите - давайте обсудим. И на Хабрахабр http://habrahabr.ru/post/146680/ текст Ралиева (28 июня 2012 в 12:44) "Современные аналоговые компьютеры: есть будущее?" и 105 комментариев к нему открывают обсуждение. Peter Belobrov 05 Apr 2015 20:24 PIT00199 Редактирование генома При чтении словосочетания "Gene Editing", которое чаще всего означает "редактирование генома" невольно возникает вопрос "а редактор кто и что и как он редактирует?" Редактирование генома, или редактирование генома инженерными нуклеазами (GEEN = ГРИН) - тип генной инженерии, в которой ДНК вставляется, заменяется или удаляется из генома с помощью искусственно сконструированных нуклеаз, или "молекулярных ножниц" (см. PIT00183). После двух встреч (вместе более 10 часов) с Л.И. Патрушевым, который руководит группой анализа и коррекции генома в лаборатории биотехнологии ИБХ РАН, идея реализации полииндикаторных чипов (пличипов) стала реальностью, которую я буду обсуждать на семинаре сегодня. Детальное изучение геномов растений, животных и человека открывает широчайшие возможности применения полученных знаний в биотехнологии и медицине. Однако только данных о нуклеотидных последовательностях геномов недостаточно для понимания функциональных взаимосвязей отдельных элементов геномов и их роли в формировании фенотипических признаков и патогенезе отдельных заболеваний. В постгеномную эпоху активно развиваются методы, позволяющие манипулировать с ДНК в геномах, а также визуализировать и управлять экспрессией генов и работой регуляторных элементов. Тем не менее далеко не все методы отвечают высоким требованиям к их эффективности, безопасности и доступности для широкого круга исследователей. В последние несколько лет появились новейшие методы редактирования геномов это системы TALEN (Transcription Activator-Like Effector Nucleases) и CRISPR (Clustered Regulatory Interspaced Short Palindromic Repeats)/Cas9. Эти появившиеся относительно недавно системы уже зарекомендовали себя как эффективные и надежные инструменты геномной инженерии. Данный обзор в основном посвящен применению указанных систем для редактирования геномов основных модельных объектов современной биологии, а также для функционального скрининга геномов, создания клеточных моделей наследственных заболеваний человека, изучения эпигеномов и визуализации клеточных процессов. Кроме того, рассмотрены основные методы конструирования подобных систем, проанализировано их действие, обсуждаются некоторые проблемы, которые ожидают исследователей при применении этих инструментов редактирования геномов. Это аннотация обзорной статьи Вопрос в начале поста пока открыт, думаю. Peter Belobrov 03 Apr 2015 01:56 PIT00190 RNA-seq etc. А мы-то умные какие!!! Только что (19/1/15) опубликованная в сети статья заставила меня передвинуть пост с кластерами воды на поздний номер, т.к. появилось совершенно неожиданное подтверждении правильности нашего подхода к родословной клеток. Жаль, конечно, что Ира бросила занятия клеточной родословной, но я точно не брошу, а после такой статьи увеличу мощность поиска новых методов изучения родословной клетки или CellBook по-нашему. Итак, коллегам из Германии и Англии пришлось вводить скрытые переменные совсем как в квантовой физике на заре её развития. И для чего? Чтобы сохранить представление о чистой клеточной линии (lineage) без перемешивания. Но с нашей точки зрение наследование и есть по сути перемешивание клеточных линий, а "чистых линий" в природе не существует, как и чистых клеточных культур, сколь бы стерильными они не были! Это преамбула. Вот статья «Вычислительный анализ поклеточной гетерогенности в данных одноклеточного РНК-секвенирования выявляет скрытые субпопуляции клеток» [Buettner2015]. Вот аннотация. Новые разработки позволили проводить беспристрастный анализ транскриптомов сотен клеток, открывая возможность для обнаружения новых субпопуляций клеток. Однако влияние потенциальных мешающих факторов, таких как клеточный цикл, на гетерогенность экспрессии генов и, следовательно, на способность надежно идентифицировать субпопуляции остается неясным. Представлен и проверен вычислительный подход, который использует модели скрытых переменных для учета таких скрытых факторов. Мы показали, что наша модель одноклеточных латентных переменных (scLVM) позволяет идентифицировать необнаружимые в противном случае субпопуляции клеток, которые соответствуют различным стадиям во время дифференциации наивных Т-клеток в 2 Т-хелперные клетки. Наш подход может быть использован не только для идентификации клеточных субпопуляций, но также для выявления различных источников гетерогенности экспрессии генов в одноклеточных транскриптомах. {Buettner2015} Florian Buettner, Kedar N. Natarajan, F. Paolo Casale, Valentina Proserpio, Antonio Scialdone, Fabian J. Theis, Sarah A. Teichmann, John C. Marioni, and Oliver Stegle. Computational analysis of cell-to-cell heterogeneity in single-cell RNA-sequencing data reveals hidden subpopulations of cells // Nature biotechnology 33 (2), 155 (2015). doi 10.1038/nbt.3102 Cited by 588 (1/8/2019) Продолжение следует PIT00470 Peter Belobrov 03 Feb 2015 02:08 PIT00174 Принципы биологии ab initio Точные ab initio биологические принципы, которые любая математическая и/или физическая теория обязана учитывать: Глава 1. Действие — это закон жизни Towards biological mathematics and physics Chapter 1. Act life is action (беспокойство) *Развернутое содержание и основная литература отдельно. (С) Peter I Belobrov, 23 Nov 2014 Peter Belobrov 03 Jan 2015 18:40 PIT00124 Сборки (assemblages) в клетке Представляю Вам новые субклеточные структуры сборки (assemblages) - функциональные блоки, образованные путем разделения клеточных фаз. Примерами сборок являются цитоплазматические P-гранулы, гранулы зародышевых клеток, и различные ядерные тельца (ядрышки, тельца Кахаля или спиральные тельца — небольшие органеллы, ассоциированные с ядрышком, тельца фактора промиелолейкоза или PML тельца, спеклы или пятнышки, etc), но эти частицы не имеют ясно объединяющей терминологии. Таким образом, предлагаемое объединяющее имя "сборки" для всех этих функциональных частиц, которые включают фазовые переходы, некоторые из них сделаны из природно неупорядоченых белков (IDPs). Эти частицы обладают поведением жидкости, так как их компоненты находятся в постоянном и быстром обмене с окружающей цитоплазмой или нуклеоплазмой. Ключевым свойством комплексов, образованных повторяющимися, с низким уровнем сложности, последовательностей IDPs является обратимость. Насколько сборка окажется универсальной субклеточной структурой - покажет будущее. А пока обратимся к очень хорошему обзору {Toretsky2014} J A Toretsky & P E Wright. Assemblages - Functional units formed by cellular phase separation // JCB, 206 (5), 579-588 (2014). DOI 10.1083/jcb.201404124 Аннотация. Разделение внутриклеточного пространство за мембранно-связанными органеллами может быть достигнуто белками, которые поливалентные, или содержат, по сути неупорядоченные области белков низкой сложности. Эти белки могут претерпеть изменение физической фазы с образованием функциональных гранулы или другие объекты в цитоплазме или нуклеоплазме, что мы всё вместе определяем термином "сборка". От природы неупорядоченные белки (Intrinsically Disordered Proteins или IDPs) играют важную роль в формировании подмножества клеточных комплексов путем стимулирования разделения фаз. Точки зрения последних работ вовлекают комплексы в болезненные состояния, указывая, что внутренние расстройства и фазовые переходы должны быть учтены при разработке терапевтических средств. Вот рисунки из статьи с полными подписями к рисункам. 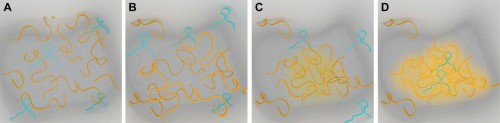 Рис. 1. [Toretsky2014] Образование сборки приводит к возникновению новых свойств белка и связывания РНК. Эта серия картинок (А-D) показывает, что увеличение локальной концентрации белка (желтые ленты) в областях клетки могут привести к фазовым переходом (желтый туман) чтобы сформировать сборку, когда достигается критическая концентрация. Фазу отделения сборки может быть образована с помощью слабых однотипных или гетеротипические взаимодействий и возможностью обмена составных молекул с окружающим раствором. Этот фазово-разделенный материал позволяет захватить и взаимодействовать с другими типами белков или РНК (голубые молекулы). (D), окончательное формирование сборки показывает поглощения двух молекул РНК. 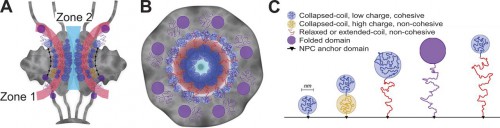 Рис. 2. [Toretsky2014] Фазовый переход вызван природно-неупорядоченными белками (IDPs) регулирует транспорт в каналах ядерных пор. (A) Сагиттальная плоскость ядерной поры, которая заполняется IDPs. FG белки, и варианты, описанные в тексте, вносят свой вклад в "начинку" пространства между структурными компонентами белка (серый цвет). Архитектура комплекса ядерной поры (NPC) создает зоны транспорта, указанные здесь как светло-красный и светло-голубой. (В) Поперечный вид сложных компонентов ядерной поры (Nups) показывает локализацию FG белков и транспортных зон. (C) Подписи определяют типы белков, обнаруженных в поровом комплексе ядра и их биофизических характеристик. Эта часть рисунка основана на Saccharomyces cerevisiae FG Nups и адаптирован из Yamada et al. (2010). 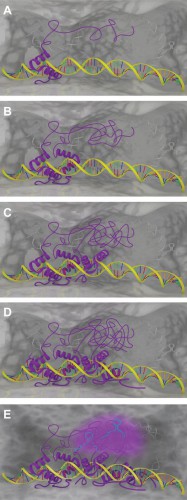 Рис. 3. [Toretsky2014] ДНК, действует в качестве каркаса для EWS-FLI1 связывания с GGAA повторами, что приводит к фазовому переходу, предположительно основанному на высокой концентрации EWS доменов. Ряд картинок (А-D) показывает последовательное связывание EWS-FLI1 (фиолетовый со спиральной областью) с GGAA (красный/зеленый) повторами в ДНК. Высокая концентрация EWS доменов происходит в результате связывания нескольких белков EWS-FLI1 в микросателлитной ДНК, что и может привести к фазовым переходом, основанных на принципиально неупорядоченных повторах с низкой сложностью. (E) Увеличилась локальная концентрация этих EWS доменных субъединиц имеют эмержентные (возникающие) свойства в критической концентрации, изображенной здесь, как пять белков, потому фазовый переход приводит к связыванию РНК (голубой). Сборка и ей взаимодействие с РНК может быть частью транскрипции или посттранскрипционной перестройки. Захват РНК может ограничивать эту динамическую фазу, отделяющую сборку от зарождающейся пре-мРНК или от посттранскрипционного комплекса сплайсинга. Сокращения - FLI1 - протоонкоген низкой сложности, (EWS) - саркома Юинга Ewing sarcoma. Читать обзор надо достаточно внимательно! Ведь он написан прямо для нашего проекта CellBook. Добавить к примерам сборок можно P-гранулы, которые несколько раз обсуждались здесь, биочастицы Ивана, другие биочастицы, гипотетические сборки Якимова (ламины в хромосомых) и целый ряд других сборок, например, биологических устройств и приводов, и даже клеточных приборов. Думается, что большие биомолекулы с растворителем также могут быть сборками. Peter Belobrov 25 Sep 2014 18:18 Проблема клеточной родословной - Cell Book (= "мусорная" ДНК ?) - в названии этого поста тесно связана с двумя предыдущими постами PIT00003, PIT00071 и PIT00072 и выражает смысл родившейся догадки в результате работы со студентами: И.В. Беляниной, В.В. Бирюковым, Е.И. Бондар и со всей нашей командой МОЛПИТ. Догадка не хилая, поэтому требует хорошие аргументы, сильные доказательства и оформления для публикации в самом "крутом" журнале. Подготовку к публикации надо делать быстро и без афиши, как учил меня автор колонки Дедала в Nature. Спасибо Вам всем за то, что Вы есть и я могу обсуждать с Вами самые глупые мысли. Хотя иногда, как позже выясняется, некоторые из идей даже и не глупы. Peter Belobrov 28 Apr 2014 17:46 Павел Захаров (СФУ) прислал новость Удалось клонировать клетки взрослых людей. Как обычно, никакой детальной информации в тексте. Гугл помог найти оригинал [Chung2014], который выкладываю с доп. инфо и графическую аннотацию 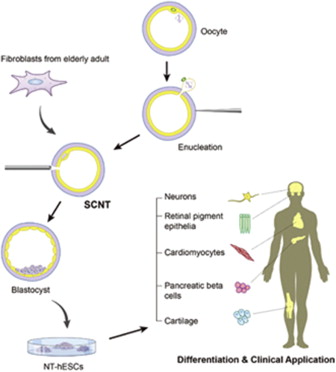 Ученым американской биотехнологической компании Advanced Cell Technology удалось клонировать клетки взрослых людей. Это открытие позволит генерировать здоровые замены для пораженных или больных клеток пациентов. Группе исследователей во главе с доктором Робертом Ланзой удалось получить стволовые клетки из тканей двух взрослых мужчин (35 и 75 лет). Процесс называется заменой ядра и предполагает взятие ДНК донора и введение его в яйцеклетку, лишенную собственной ДНК. В результате образуется единое целое и клетки начинают делиться. Дальше стволовые клетки продолжают выращивать в лаборатории. Ранее ученым удавалось клонировать стволовые клетки, взятые у эмбрионов и новорожденных. {Chung2014} YG Chung et al. Human somatic cell nuclear transfer using adult cells // Cell stem cell 14 (6), 777-780 (2014). doi 10.1016/j.stem.2014.03.015 Peter Belobrov 18 Apr 2014 19:54 Это название статьи в журнале BioTechniques сегодня привело меня в шок. Моё понимание того, что транскриптом и малые РНК - это важно, неожиданно связалось с родословной клеток. Приведу перевод (быстрый с Гуглом) фрагмента 04/16/2014 Kayt Sukel  Когда Йошихиде Хаясидзаки, директор научного центра "Oмики" (OSC) в престижном Института RIKEN в Японии, помог основать проект FANTOM (функциональная аннотация генома млекопитающих) в 2000 году, он не ожидал, что работа международного консорциума может поколебать сложившуюся догму генетической теории. ФАНТОМ появилась на свет в том же году, как и проект генома человека (HGP), и стала публичным "черновиком" генома человека. Было бы еще три года, прежде чем полная карта группа HPG был опубликован. В то время, Хаясидзаки и его коллеги хотели ФАНТОМ дополнить работу HPG, поэтому они выбрали конкретную цель: обеспечить более глубокое понимание транскриптома, сбор коллекции молекул рибонуклеиновой кислоты (РНК), в том числе, информационной РНК (мРНК), транспортная РНК (тРНК) и других РНК, создаваемых клетками. Многие группы, анализирующие транскриптом, ограничивали свое внимание только на белок-кодирующих мРНК. Доминирующая теория в области в то время был, что это те РНК, которые только и имеют смысл. Но в 2010 году, группа ФАНТОМ сделал "крутое" заявление, когда после секвенирования как кодирующих так и некодирующих белки полнометражных комплементарных ДНК (кДНК) они сообщили, что более половины генома фактически транскрибируют РНК, в том числе около 15 тысяч некодирующих белки РНК (нкРНК). "Мы обнаружили, что есть "тонны" некодирующих РНК, "тонны" из них в клетке. Это была такая интригующая находка, которая это не вписывалась в общие знания в то время", - сказал Хаясидзаки. "Это шокировало многих людей, и даже нас". Хотя они и были озадачены тем, что они нашли, группа Хаясидзаки спрашивает, играют ли эти нкРНК какую-то важную регуляторную роль в экспрессии генов. Сегодня, во многом благодаря новаторской работе Fantom, всеми признается, что более 90% генома транскрибируется. И последующие исследования Fantom показала, что интуиция Хаясидзаки попала в цель: многие нкРНК играют важнейшую роль в регуляции генов, как промоторов генов и усилителей (энхансеров) - в конечном итоге помогают направлять дифференцировку клеток. Подробнее читайте в двух статьях и в диссертации Robin Andersson et al. An atlas of active enhancers across human cell types and tissues // Nature 507, 455–461 (27 March 2014) The FANTOM Consortium and the RIKEN PMI and CLST (DGT). A promoter-level mammalian expression atlas // Nature 507, 462–470 (27 March 2014). Прикол: Scholar Google цитирует как автора (сокращает) в TF Consortium !!! Michelle Rönnerblad. Dissecting the epigenetic landscapes of hematopoiesis and fission yeast. (Прохождение эпигенетических ландшафтов кроветворения и делящихся дрожжей.) Stockholm 2014 Peter Belobrov 17 Apr 2014 14:20 Размышляя над вопросом Иры Беляниной: "т.е я могу говорить о мицелле, как о тотипотентной, из которой могут получиться разные типы...?", подумал о том, что может быть и ответ на вопрос: "где находится родословная клеток?" также прост и очевиден: в структуре хроматина! Сейчас перенес это из поста PIT00069 Peter Belobrov 28 Mar 2014 15:24 PIT00003 P гранулы Вариант 2 P гранулы или «следствие ведут колобки»ПИ Белобров & MOLPIT О сколько нам открытий чудных План статьи (После завершения план становится содержанием текста эссе, реферата, курсовой …) Abstract (аннотация). Установлено, что название «P» гранул имеет точное происхождение – “polar”! Важный результат поиска колобков состоит в понимании того, что есть ещё неоткрытые компартменты клеток. Рождение этого понятия имеет глубокий биологический смысл и связано с асимметричным делением клеток в мейозе. Введение. На семинаре МОЛПИТ 20/10/12 возник вопрос: «что означает сокращение P гранулы?» Два предположения protein и physiological были высказаны немедленно. Однако, при попытке найти в литературе первую статью, где было введено это сокращение, увеличивали число претендентов, например: perinuclear, plasmatic, polar, polarity, peripheral, posterior, proximal и т.д. Но так и не было полной ясности (точной доказательности). Методы поиска становились всё более изощрённые (часть из них будет описана в разделе материалы и методы), но результат оказался достаточно простым и глубоким одновременно. При обсуждении в МОЛПИТ 25/10/12 было несколько новых моментов отмечено: Рисунок из работы [bpq210], который Ренат поделил пополам и послужил основой: постановки задачи его курсовой работы и мысли уточнения «ажурных фаз» и дробных чисел Авогадро NAd. Четное или нечетное само число Авогадро (здесь работает принцип точного определения его: ссылка Lide + inet). Здесь биологическая математика в действии! Лучше всего название проблемы в качестве названия главы, хотя смысл содержания главы – это обзор литературы по проблеме с обязательным переходом к точной постановке задачи как части этой проблемы. TEXT IN THE PROGRESS Литература Lectures. Anthony Hyman Part 4. Formation of P granules http://www.ibioseminars.org/lectures/bio-mechanisms/anthony-hyman/anthony-hyman-part-4.html Wiki Info: http://en.wikipedia.org/wiki/Germ http://ru.wikipedia.org/wiki/Germ_(значения) http://en.wikipedia.org/wiki/Germ_cell http://ru.wikipedia.org/wiki/Гоноцит germ - (смысловая "клякса" этого слова непростая) Новые слова Gingerbread - (смысловая "клякса" этого слова такая) Peter Belobrov 31 Oct 2012 01:00
© International Open Laboratory for Advanced Science and Technology — MOLPIT, 2009–2026
|
